Class
Subclass
Order
Family
Genus
Terrestrial or rupestral ferns. Rhizomes short and erect or short-creeping, bearing scales. Rhizome scales ovate, 8–16 mm long, 1–4 mm wide, dark brown, clathrate. Fronds 105–1080 mm long. Stipes 30–540 mm long, brown abaxially and green adaxially or rarely brown throughout, bearing ovate or narrowly ovate scales with filiform apices. Rachises green, very narrowly winged in distal half, scaly. Laminae 2-pinnate in small specimens, usually 2-pinnate-pinnatifid to 3-pinnate-pinnatifid, ovate or elliptic, tapering gradually to the apex, 60–800 mm long, 24–350 mm wide, dark green on both surfaces, herbaceous, bearing scales, lacking hairs, occasionally bearing bulbils on adaxial surface. Primary pinnae in 6–26 pairs below pinnatifid apex, usually not overlapping or rarely overlapping; proximal pinnae and those at mid-lamina ovate or narrowly ovate; the longest at or below the middle, 13–225 mm long, 7–80 mm wide, apices acute to acuminate or rarely obtuse, bases stalked; costae of primary pinnae very narrowly winged, or lacking wings. Secondary pinnae gradually decreasing in length along each primary pinna to distal end, the basal acroscopic secondary pinna the largest; the longest proximal secondary pinnae narrowly ovate or ovate, 4–60 mm long, 2–25 mm wide, apices acute or obtuse, usually partly or completely divided into tertiary segments or rarely undivided, bases stalked; the distal secondary pinnae elliptic or oblong, apices acute or obtuse, bases adnate. Tertiary segments narrowly oblong or oblong or elliptic, 5–15 mm long, 0.8–7 mm wide, margins entire or divided. Sori away from margins or rarely submarginal; indusia 2–4 mm long, straight; free margins of indusia entire. Mean spore size 41–48 μm long, 28–32 μm wide; perispores irregularly ridged.
Asplenium gracillimum is a very variable species; at one extreme there are fronds with broad secondary pinnae that are divided into tertiary segments 2–7 mm wide, and at the other there are fronds that have the secondary pinnae completely divided into narrow ultimate segments 1 mm or less in width. The latter forms were referred to A. bulbiferum var. tripinnatum Hook.f. by many earlier authors, but, as noted by Brownsey (1977b), the type of Hooker’s variety is something different. The specimen is immature and its identity remains uncertain; it is included here in the synonymy for A. gracillimum but could possibly be a hybrid with A. appendiculatum. The two forms of A. gracillimum frequently grow together; the highly divided fronds are, on average, slightly smaller than the broader forms, but their overall size range falls within that for the broad forms. Genetically, several subgroups can be recognised within A. gracillimum (Perrie et al. 2010), but the highly divided forms occur in several of these and collectively they do not comprise a separate lineage. Hence the two forms are not recognised here taxonomically.
Asplenium gracillimum is often confused with both A. bulbiferum and A. hookerianum, species that are now known to be its tetraploid parents (Shepherd et al. 2008). Asplenium bulbiferum is generally a larger plant than A. gracillimum and tends to grow in wetter areas. However, the two species frequently occur together and are known to hybridise (Brownsey 1977a). Asplenium bulbiferum usually has more pronounced wings on the rachis and primary costae than A. gracillimum, and only a few of the proximal secondary pinnae on each primary pinna have distinct stalks; in A. gracillimum the secondary pinnae are usually stalked along at least half of the primary pinna. The scales of the two species are markedly different: those in A. bulbiferum are ovate and lack filiform apices, whereas in A. gracillimum they are narrower and almost always drawn out into filiform apices. In New Zealand Asplenium bulbiferum is usually bulbiferous with at least a few bulbils on the lamina, whereas A. gracillimum often lacks bulbils or is only sparingly bulbiferous. Asplenium gracillimum also has larger spores than A. bulbiferum (41–48 × 28–32 cf. 34–37 × 23–26 μm), and the two species can be reliably distinguished on this basis.
Asplenium gracillimum generally has much larger fronds than A. hookerianum, but small plants of A. gracillimum are sometimes difficult to discriminate. Usually the secondary pinnae of A. hookerianum are more regularly stalked than in very small fronds of A. gracillimum. The basal pair of pinnae are angled acroscopically, whereas those in A. gracillimum arise at right angles to the rachis, or are angled basiscopically. Spore size will also distinguish the two species; A. gracillimum has larger spores than A. hookerianum (41–48 × 28–32 cf. 31–37 × 23–27 μm).
Asplenium gracillimum can sometimes be confused with A. appendiculatum subsp. appendiculatum, but the latter has more coriaceous fronds, always lacks bulbils, has primary pinnae with longer acuminate apices, and has longer indusia (2–8 mm cf. 2–4 mm long). Highly divided forms of A. gracillimum can appear very similar to A. richardii when growing at higher altitudes, but the scales of the latter species are rather longer and narrower than in A. gracillimum.
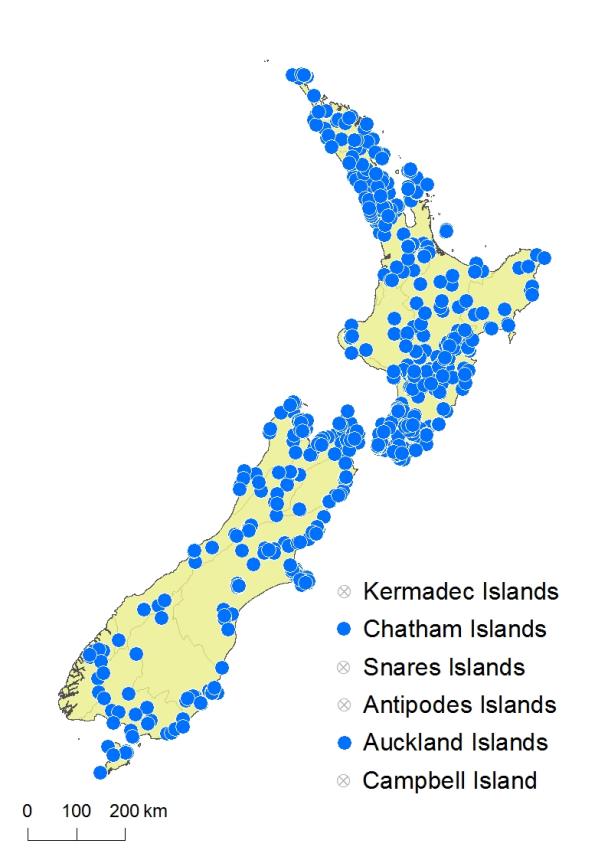
North Island: Northland, Auckland, Volcanic Plateau, Gisborne, Taranaki, Southern North Island.
South Island: Western Nelson, Sounds-Nelson, Marlborough, Westland, Canterbury, Otago, Southland, Fiordland.
Chatham Islands, Stewart Island, Auckland Islands.
Altitudinal range: 0–1000 m.
Asplenium gracillimum occurs throughout coastal, lowland, and montane areas of the North Island from Te Paki southwards. It extends from near sea level to over 1000 m in the Ruahine Ranges. In the South Island it grows widely in lowland and montane areas except for the driest parts of inland Otago, Canterbury and Marlborough. It ranges from near sea level to 900 m at Jordan Stream, Marlborough. It extends to the Chatham Islands, Stewart Island, and Auckland Islands.
Also Australia (Queensland, New South Wales, Victoria, Tasmania).
A terrestrial fern that occurs under kauri, podocarp, broadleaved, and beech forest, and under tall kānuka forest. It grows on drier well-drained soils on the forest floor, often on steep slopes, on dry ridges, in gullies, amongst rocks or tree roots, on banks, on logs, by streamsides, on river terraces, in shaded road cuttings, and under rock overhangs. Sometimes it is found in reverting pasture or grazed bush, on stabilised dunes under scrub, or under Pinus radiata. It occurs on a variety of rock types, including greywacke, sandstone, schist, limestone, and marble.
There is evidence that A. gracillimum hybridises with A. bulbiferum, A. hookerianum, A. obtusatum (Brownsey 1977a), A. appendiculatum subsp. appendiculatum (Brownsey 1983), A. flaccidum subsp. flaccidum, A. lyallii (de Lange et al. 2011), A. appendiculatum subsp. maritimum and A. richardii (newly recorded here).
n = 144 (Brownsey 1977b).
Typification of Asplenium gracillimum has proved to be difficult. Allan (1961, p. 72) stated that "The type in W[ELT] (Dannevirke. W.C.) is one complete plant" and cited a series of measurements taken from the fronds. However, there are numerous sheets of A. gracillimum in the Colenso Herbarium at WELT; none of them can be unambiguously linked to Allan’s description, and hence his statement does not constitute lectotypification. Brownsey (1977b) identified WELT P003348 as the holotype. However, he later noted that more material had been found (Brownsey 1998), and a series of eight sheets was examined. These proved to be a mixture of A. gracillimum and hybrids between it and A. hookerianum. WELT P003348 was shown to be one of the hybrid plants, and a lectotype (WELT P002858) was selected consistent with the retention of the name A. gracillimum for the species rather than a hybrid.
DNA analyses strongly suggest that A. gracillimum is an allopolyploid species derived from A. bulbiferum and A. hookerianum, with multiple independent polyploid origins (Shepherd et al. 2008; Perrie et al. 2010).