Class
Subclass
Order
Family
Genus
Terrestrial or rupestral ferns. Rhizomes short and erect or short-creeping, often forming a hard woody mass above ground, bearing scales. Rhizome scales narrowly ovate, 10–25 mm long, 1–5 mm wide, brown, clathrate. Fronds 35–640 or rarely up to 790 mm long in sheltered sites. Stipes 10–270 or rarely up to 320 mm long, dark brown proximally, green distally, bearing narrowly ovate or narrowly triangular scales with short filiform apices. Rachises green, scaly. Laminae 1-pinnate, ovate or elliptic, narrowed to a short undivided or rarely lobed terminal segment with serrate margins similar to the primary pinnae; laminae 23–370 or rarely up to 470 mm long, 15–160 or rarely up to 210 mm wide, dull green on both surfaces, thick and fleshy, bearing scales on abaxial surface, lacking hairs. Primary pinnae in 3–28 pairs below an undivided apical segment, not or only slightly overlapping; proximal pinnae ovate or elliptic, those at mid-lamina ovate or elliptic or oblong; the longest pinnae at or below the middle, 7–90 or rarely up to 105 mm long, 5–25 mm wide; pinna apices obtuse or rounded or rarely acute, margins serrate, bases stalked. Sori away from margins; indusia 2–11 mm long, straight; free margins of indusia entire. Mean spore size 45–52 μm long, 29–33 μm wide; perispores slightly winged but with few crests or ridges.
Asplenium decurrens is morphologically very similar to A. obtusatum and A. oblongifolium. It occupies a similar coastal habitat to A. obtusatum and is only distinguished by its slightly shorter and narrower laminae (23–370 mm long, 15–160 mm wide, cf. 27–540 mm long, 23–290 mm wide), narrower stipe scales, larger spores, and octoploid chromosome number. However, the two species are geographically separated, A. decurrens occurring north from the north Taranaki coast, and A. obtusatum only south from Kapiti Island. Asplenium decurrens is distinguished from A. oblongifolium by its thick, fleshy fronds, rather round-ended pinnae, and narrowly ovate rather than hair-like stipe scales. Both species can occupy coastal habitats, but A. decurrens does not extend into inland areas.
Plants of A. decurrens produce their largest fronds on the Kermadec and Three Kings Islands. They gradually diminish in size southwards, with only rather stunted plants occurring around Auckland and the western Waikato. This is in contrast to A. obtusatum, which produces its largest fronds in the subantarctic and gradually decreases in size northwards.
North Island: Northland, Auckland, Volcanic Plateau, Gisborne, Taranaki.
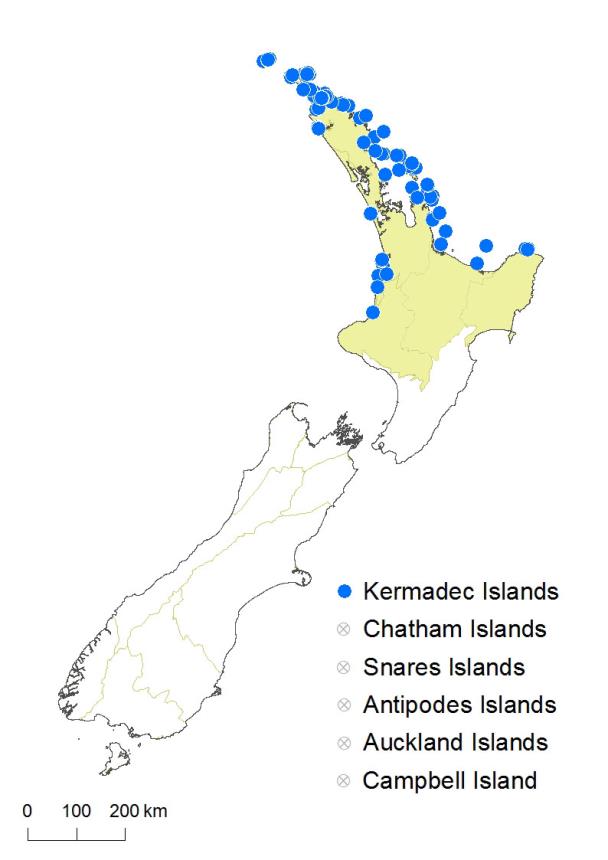
Kermadec Islands, Three Kings Islands.
Altitudinal range: 0–200 m.
Asplenium decurrens occurs on the northern and southern Kermadec Islands (Sykes 1977; de Lange 2015a, 2015b), the Three Kings Islands, and in the northern part of the North Island. On the west coast there are isolated records from Mitimiti and Karekare, with more extensive populations from Kāwhia Harbour to Tongapōrutu in north Taranaki. On the east coast, the species extends from Te Paki along the east coast of Northland and the islands of the Hauraki Gulf to Coromandel and the Bay of Plenty to near East Cape. It is confined to coastal regions, ranging from sea level to 160 m on sea cliffs at Mt Karioi and over 200 m on Macauley Island in the Kermadec group.
Octoploid populations of A. decurrens are believed to occur in Australia, based on spore size (Brownsey 1998). Morphologically similar plants, which are probably also A. decurrens, are known from some Pacific islands (New Caledonia, Tonga, French Polynesia, Pitcairn and Easter Island), but their identity needs confirmation.
Asplenium decurrens grows on exposed coastal rocks and cliffs, in crevices and amongst boulders, and in low coastal scrub. It is found on a variety of rock substrates, including andesite, basalt, breccia, greywacke, lava, limestone, calcareous mudstone, sandstone, scoria, pumice, and guano.
There is evidence that A. decurrens (as A. obtusatum subsp. northlandicum) hybridises with A. oblongifolium, A. shuttleworthianum (Brownsey 1977a), and A. flaccidum subsp. haurakiense (newly recorded here).
n = 144 (Brownsey 1977b – as A. obtusatum subsp. northlandicum).
The new combination Asplenium northlandicum (Brownsey) Ogle was published in Vol. 25, No. 4, of the NZ Journal of Botany. Although dated 1987, it was not published until April 1988, as indicated in Vol. 26, No. 1.
Asplenium decurrens was previously treated as A. obtusatum subsp. northlandicum, an octoploid subspecies (Brownsey 1977b) suggested to be an autopolyploid derivative of the tetraploid A. obtusatum (Brownsey 1977a). However, Shepherd et al. (2008) used nuclear DNA sequence data from New Zealand material to show that this origin is unlikely. Rather, they suggested that A. decurrens (as A. northlandicum) is an allopolyploid, with either A. obtusatum or A. oblongifolium as the chloroplast parent and an unknown tetraploid as the other parent.
That conclusion supported the recognition of the octoploid as a distinct species, rather than as a subspecies of the tetraploid A. obtusatum. Ogle (1988) had previously raised the subspecies to species level, as A. northlandicum, but at this rank there are actually two earlier available names when Australian and New Zealand populations are treated as the same taxon – A. decurrens Willd. and A. sarmentosum Willd. (Brownsey & Perrie 2016). The two species were described by Willdenow (1810) from Tasmania and distinguished from A. obtusatum G.Forst. and A. obliquum G.Forst., which Willdenow believed to be confined to New Zealand. Earlier, Labillardière (1807) had accepted A. obtusatum and A. obliquum in Australia as well as New Zealand. Brownsey (1977a) reduced A. obliquum to synonymy with A. obtusatum, accepting the latter name as the earliest available for the tetraploid species. It is present in New Zealand but not in Australia. However, the octoploid cytotype is now believed to occur in both New Zealand and Australia, and when treated at species level, either of Willdenow’s names could be applied to it. Asplenium decurrens was preferred by Brownsey & Perrie (2016) because Willdenow indicated that he had seen a specimen of it, and the holotype collected by Labillardière is extant in the Willdenow herbarium (B-W 19885-01 0). In contrast, Willdenow gave no indication of having seen a specimen of A. sarmentosum, none are now extant in the Willdenow herbarium, and the holotype of A. sarmentosum must therefore be the illustration in Novae Hollandiae Plantarum Specimen (Labillardière 1806–1807) cited by Willdenow (1810).