Class
Subclass
Order
Family
Genus
Terrestrial or rupestral ferns. Rhizomes short, erect, bearing scales. Rhizome scales acicular with short apices, 2–5 mm long, 0.2–0.5 mm wide, dark brown, clathrate with an occluded central midrib. Fronds 25–300 mm long. Stipes 2–55 mm long or rarely to 80 mm long, dark brown, very narrowly winged to the base, shiny, stiff, polished, glabrous except at very base. Rachises dark brown, very narrowly winged throughout, glabrous. Laminae 1-pinnate, linear, tapering gradually to the apex, 25–290 mm long, 4–21 mm wide, dark green on both surfaces, coriaceous, glabrous. Primary pinnae in 10–36 pairs below a small apical segment, not overlapping, all oblong or broadly elliptic or broadly ovate except 1–2 proximal pairs often greatly reduced and widely spaced; the longest at about the middle, 2–10 mm long, 1–7 mm wide; pinna apices obtuse or rounded, margins shallowly crenate or serrate, bases cuneate or acute, sessile or very shortly stalked or rarely 1–2 proximal pairs adnate. Sori away from margin; indusia 1–3 mm long, straight; free margins of indusia irregularly incised. Mean spore size 34–47 μm long, 31–33 μm wide; perispores with numerous crests and ridges.
Asplenium trichomanes is recognised by its dark brown polished stipes and rachises, narrow pinnate fronds, and sessile, oblong, or ovate pinnae. It is similar to A. flabellifolium but readily distinguished by its brown, rather than green, stipes and rachises. It also lacks the characteristic rachises of A. flabellifolium that extend beyond the distal pinnae and root at their tips. Asplenium flabellifolium typically also has flabellate pinnae, while the pinnae of A. trichomanes are oblong to ovate.
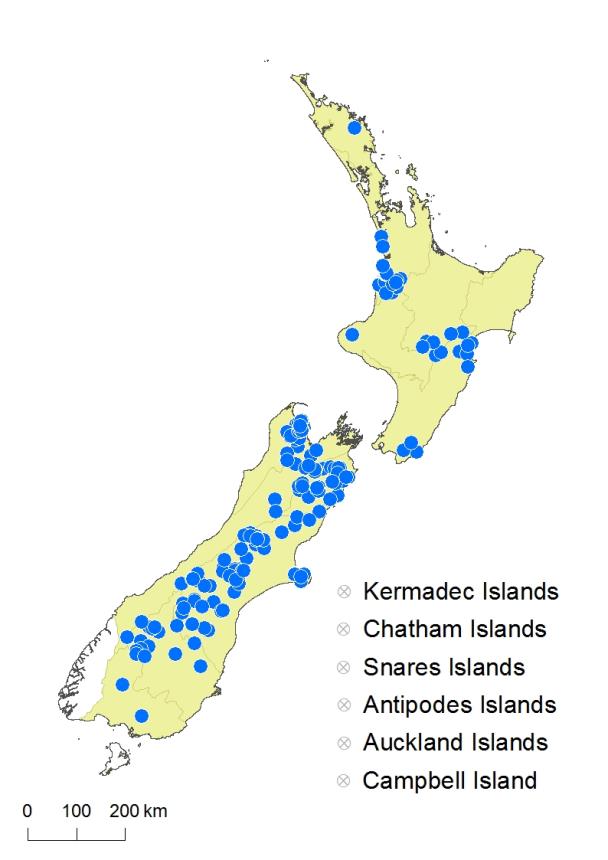
North Island: Northland, Auckland, Volcanic Plateau, Gisborne, Taranaki, Southern North Island.
South Island: Western Nelson, Sounds-Nelson, Marlborough, Westland, Canterbury, Otago, Southland.
Altitudinal range: 5–1375 m.
Asplenium trichomanes has a scattered distribution in the North Island, where it occurs mainly on limestone; populations are known at Kawakawa, Northland, from Port Waikato to Waitomo, from Hawke’s Bay to the Ruahine Ranges, and in southern Wairarapa. Cheeseman (1906) also recorded this species from Mt Taranaki, but there is no specimen in his herbarium. There is a single collection from this location without collector or date (AK 114914), but it has not been collected there for over a century and needs confirmation. The species occurs in coastal and lowland areas from near sea level, up to 1000 m in the Kaimanawa Mountains. In the South Island it grows in lowland to montane areas extending locally to the sub-alpine, mainly on the eastern side of the island from Nelson to the Hokonui Mountains, Southland. It ranges from 50 m at Tākaka to 1375 m on Mt Owen, north-west Nelson, and in the Inland Kaikōura Range. It is not known from Fiordland, Stewart Island, or the Chatham Islands.
Also North America, Europe, Asia, Africa, Australia (Western Australia, South Australia, Queensland, New South Wales, Victoria, Tasmania).
Asplenium trichomanes grows most frequently in crevices and fissures of limestone or marble, but also occurs on greywacke, schist, and serpentine. It grows on open exposed rocks, bluffs, cuttings, and banks, under overhangs, or on rock in the shade of podocarp, broadleaved or beech forest, kānuka or mānuka, or under scrub. Occasionally it extends into sub-alpine tussock, and around Kāwhia Harbour it grows close to the sea.
Asplenium trichomanes is unusual among New Zealand species of the genus in that it has not been recorded hybridising with any other species. Its isolated phylogenetic position in Clade XI of Ohlsen et al. (2014), in contrast to most New Zealand species, which occur in Clade V, may account for this.
n = 72, 108 (Brownlie 1954; Dawson et al. 2000).
Two chromosome races, representing tetraploid and hexaploid forms, have been reported for A. trichomanes in New Zealand (Brownlie 1954; Lovis 1956, 1958; Dawson et al. 2000). Lovis (1956, 1958) noted that the hexaploid could only be distinguished morphologically by the nature of the rhizome scales, which have a broader occluded central midrib than in the tetraploid, and by the size of the spores (34–39 μm long in the tetraploids and 39–47 μm in the hexaploids). Only three collections of the tetraploid were known to Lovis, none of them now represented in New Zealand herbaria, but stated to have come from Tangoio, north-east of Napier, from Pukeora Sanatorium, west of Waipukurau, and from the upper reaches of the Tukipo River.
The two cytotypes probably deserve recognition as separate taxa, but whether at specific or subspecific level requires further investigation. The correct names are also in doubt. The hexaploid was described as A. melanolepis by Colenso (1888), but this name is illegitimate because of an earlier homonym, and the taxon requires a new name at either specific or subspecific rank.
The diploid taxon in Europe has traditionally been known as A. trichomanes L. or A. trichomanes subsp. trichomanes (Lovis 1964), and the tetraploid taxon as either A. trichomanes subsp. quadrivalens D.E.Mey. or A. quadrivalens (D.E.Mey.) Landolt (Meyer 1962; Landolt 2010). A lectotype for A. trichomanes L. was designated in this sense by Viane (in Jonsell & Jarvis 1994), but his selection is pre-dated by Bobrov (1984), who designated a tetraploid specimen in LINN as the lectotype (see Lin & Viane 2013). If confirmed, Linnaeus’s name would therefore apply to the tetraploid taxon and not to the diploid. Lin & Viane (2013) indicated that a proposal might be made to conserve the name A. trichomanes sensu stricto with a conserved type. Pending such a proposal, we use the name A. trichomanes here in a broad sense to encompass both the cytotypes present in New Zealand.
The tetraploid taxon is widely distributed in North America, Europe, Asia, Africa and Australia (Western Australia, South Australia, New South Wales, Victoria, Tasmania), but the hexaploid is known only from New Zealand, apart from a single collection from ACT, Australia (Brownsey 1977, 1998).
The tetraploid taxon was given a conservation status of Nationally Critical by de Lange et al. (2013).