Class
Subclass
Order
Family
Genus
Rhizomes erect, up to 70 mm long (in herbarium material), bearing scales. Rhizome scales linear to narrowly ovate, 5–12 mm long, 0.4–0.7 mm wide, dark brown, concolorous. Fronds strongly dimorphic; sterile fronds 120–700 mm long, forming a prostrate to suberect rosette; fertile fronds on same plant usually a little shorter, 85–620 mm long, positioned centrally, held erect. Sterile fronds. Stipes 10–140 mm long, red-brown, bearing a few scales proximally. Rachises yellow-brown, grooved adaxially, glabrous. Laminae 80–600 mm long, 15–115 mm wide, or very rarely to 125 mm wide, deeply pinnatifid to pinnatisect, narrowly elliptic to narrowly obovate, with a short apical segment, similar colour on both surfaces, red-tinged when young, herbaceous to coriaceous, glabrous. Pinnae in 7–54 pairs, closely spaced, narrowly oblong or narrowly triangular or triangular, straight or falcate, gradually reducing to short, rounded flanges at the lamina base; the longest pinnae at or above the middle, 14–60 mm long, or very rarely only 8 mm long or up to 68 mm long, 6–15 mm wide, or rarely only 4 mm wide, apices usually acute or rarely obtuse, margins shallowly crenate, bases adnate; the basal pinnae sub-opposite or alternate. Veins free. Fertile fronds. Stipes 20–250 mm long. Laminae 40–530 mm long, 10–70 mm wide. Pinnae in 15–52 pairs, linear, straight or falcate, 5–40 mm long, 1–3 mm wide, reducing to tiny sterile flanges at the lamina base. Sori and indusia in one row either side of costa, continuous along the length of the pinna.
Blechnum chambersii is recognised by its dimorphic fronds, free veins, pinnatisect sterile laminae, and continuous sori. The sterile laminae are narrowly elliptic to narrowly obovate, with oblong to almost triangular pinnae that are usually acute at their apices, shallowly crenate on the margins, and adnate at their bases. The basal pinnae are alternate or sub-opposite. The laminae are glabrous and a similar colour on both surfaces. The sterile fronds form a prostrate to suberect rosette with the fertile fronds held upright in the centre, the fertile usually a little shorter than the sterile.
Blechnum chambersii is very similar to B. membranaceum, the two species hybridising in areas of overlap, but plants of B. chambersii can be distinguished by their generally broader laminae (15–115 mm wide, cf. 8–40 mm wide), elliptic rather than obovate outline, shallowly toothed pinnae, which are alternate at the base of the lamina rather than in opposite pairs, and by their fertile fronds which are a little shorter, not longer, than the sterile fronds. B. chambersii is also similar to B. banksii but can be distinguished by its sterile laminae, which are generally broader (15–115 mm wide, cf. 5–40 mm wide) and never fleshy.
Blechnum norfolkianum is closely related to B. chambersii, differing principally in its larger fronds, wider laminae, shiny green colour, more acuminate pinna apices, and oblong basal flanges. In B. chambersii the sterile laminae are 15–115 mm wide and the longest pinnae 14–60 mm long, compared to 70–250 mm wide and 50–125 mm long in B. norfolkianum. The fertile laminae are 10–70 mm wide and the longest pinnae 5–40 mm long in B. chambersii, and 50–190 mm wide and 25–100 mm long in B. norfolkianum. The lamina length:width ratio is c. 3–5:1 in sterile fronds and c. 2–5:1 in fertile fronds of B. norfolkianum compared to ratios of c. 4–6.5:1 and c. 5–10:1 respectively in B. chambersii. Very large fronds of B. chambersii resembling B. norfolkianum are found in some places between Auckland and Tauranga, but they can usually be distinguished by the length:width ratio of the laminae.
Occasional aberrant forms of B. chambersii are found, including fronds with very long, narrow laminae, sterile laminae with occasional fertile pinnae, and crested forms.
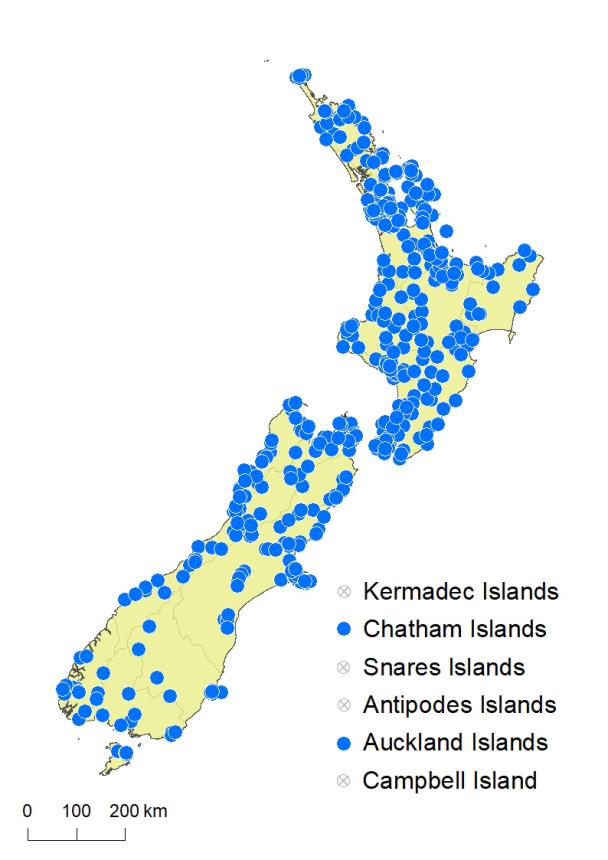
North Island: Northland, Auckland, Volcanic Plateau, Gisborne, Taranaki, Southern North Island.
South Island: Western Nelson, Sounds-Nelson, Marlborough, Westland, Canterbury, Otago, Southland, Fiordland.
Chatham Islands, Stewart Island, Auckland Islands.
Altitudinal range: 0–1100 m.
Blechnum chambersii occurs in lowland and montane areas throughout the North Island from Te Paki to Wellington. It grows from near sea level, up to 1100 m in the Kaimanawa Ranges. In the South Island it occurs in lowland and montane regions throughout, except for the drier parts of Otago and south Canterbury, reaching 1100 m near Lake Shirley, Fiordland. It also extends to the Chatham Islands and Stewart Island. Plants on the Chatham Islands are generally larger; a few approach B. norfolkianum and require further investigation. It was recorded for the Auckland Islands by Johnson & Campbell (1975 – OTA 33135, 33136), and has also been observed there (see iNaturalist 38523320), but no collections are held in AK, CHR or WELT. Further collections are desirable to confirm its presence on the Auckland Islands.
Also Australia (South Australia, New South Wales, Victoria, Tasmania).
Blechnum chambersii is a terrestrial fern found under kauri, podocarp, broadleaved or beech forest, under mānuka, kānuka and coastal scrub, and under Salix species in swampy areas. It grows in rock crevices, under overhangs, on damp rock walls, near waterfalls, on rocky banks and roadside banks, on the forest floor, on stream banks in wet gullies, and rarely as a low epiphyte, usually favouring shaded habitats.
There is evidence for hybridisation between Blechnum chambersii and B. membranaceum (see Blechnum ×aggregatum; Tindale 1960).
It is likely that B. chambersii hybridises with B. norfolkianum on offshore islands where the two species often occur together, and where they are sometimes hard to distinguish. However, no sign of spore abortion has been seen in plants of intermediate morphology, and conclusive evidence of hybridisation is lacking.
2n = 66 (Chambers 1954 – as B. lanceolatum).
Allan (1961) treated this species as B. lanceolatum (R.Br.) J.W.Sturm, but that name is pre-occupied by B. lanceolatum Raddi. When treated in Austroblechnum, the correct name is A. lanceolatum.
DNA sequences for a single chloroplast locus from New Zealand Blechnum chambersii and B. norfolkianum were found to be identical, whereas one of two B. membranaceum samples differed by one base pair (Shepherd et al. 2007). Wilson (2012), using DNA sequences from three chloroplast loci, found samples of B. membranaceum and B. norfolkianum from New Zealand, and Australian and New Zealand B. chambersii, to differ by up to seven base pairs, but with no strongly supported relationships among them. The two studies indicate that all three taxa are very closely related.
B. chambersii has been reported from Fiji (Brownlie 1977, as Blechnum doodioides (Brack.) Brownlie; Nakamura 2008; Brownsey & Perrie 2011), but plants from there have much wider fertile fronds, as well as DNA sequence differences (Perrie et al. 2014). It is likely that they belong to a different species.