Class
Family
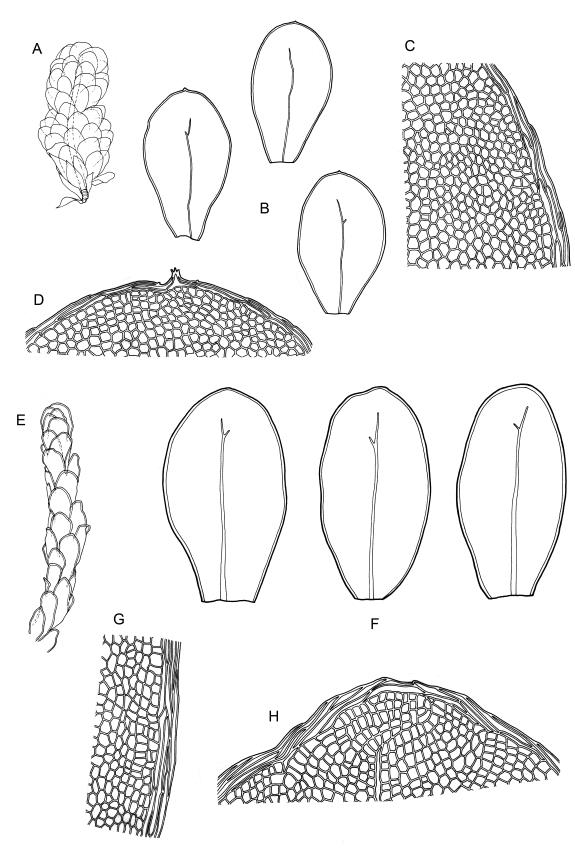
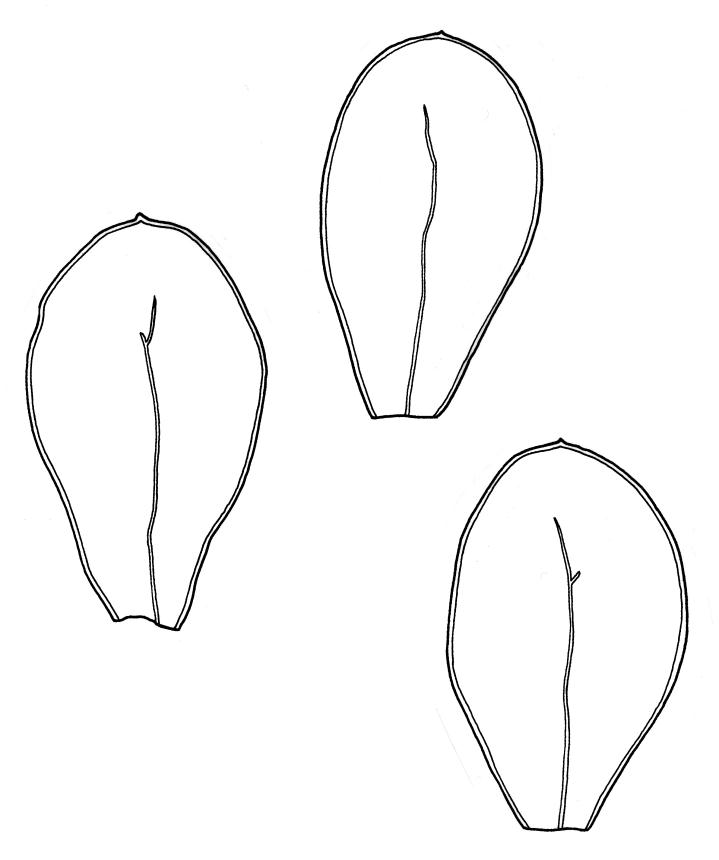
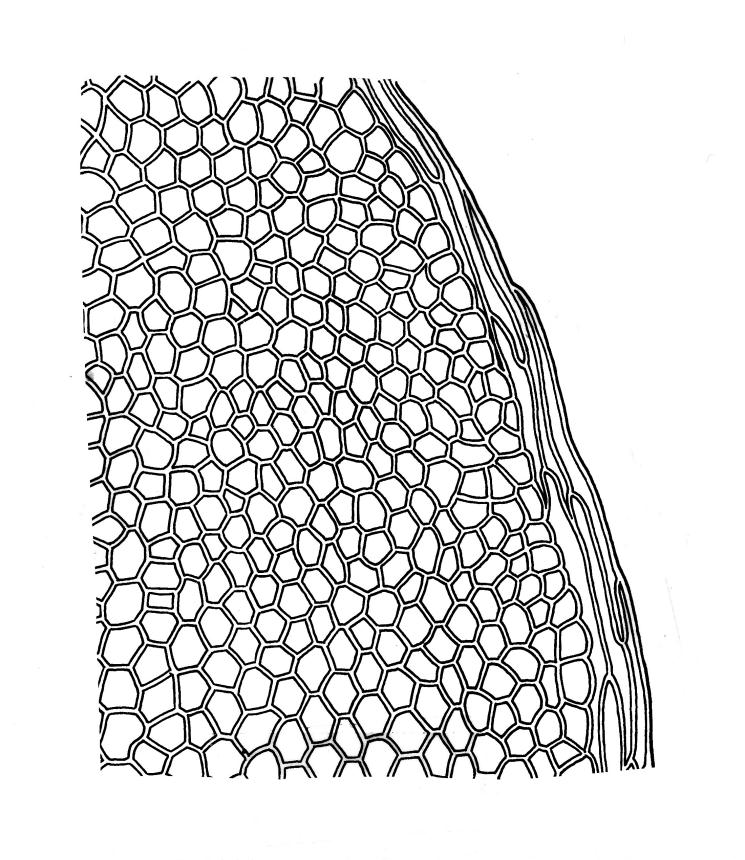
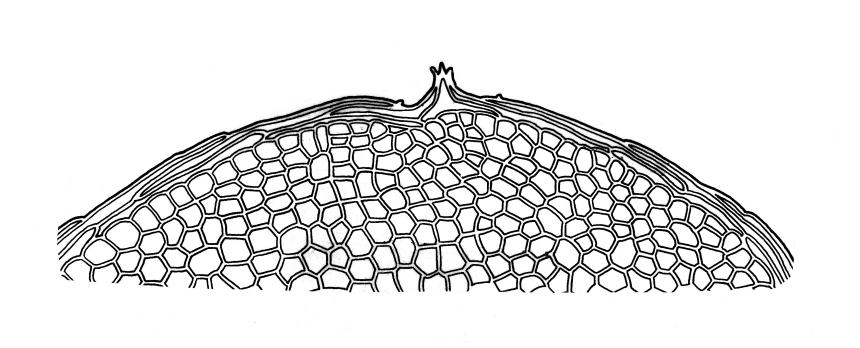
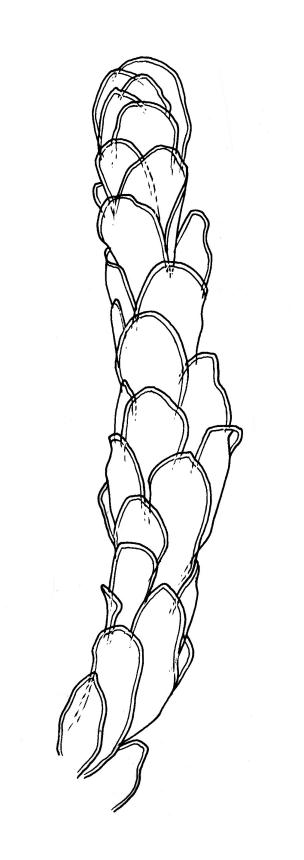
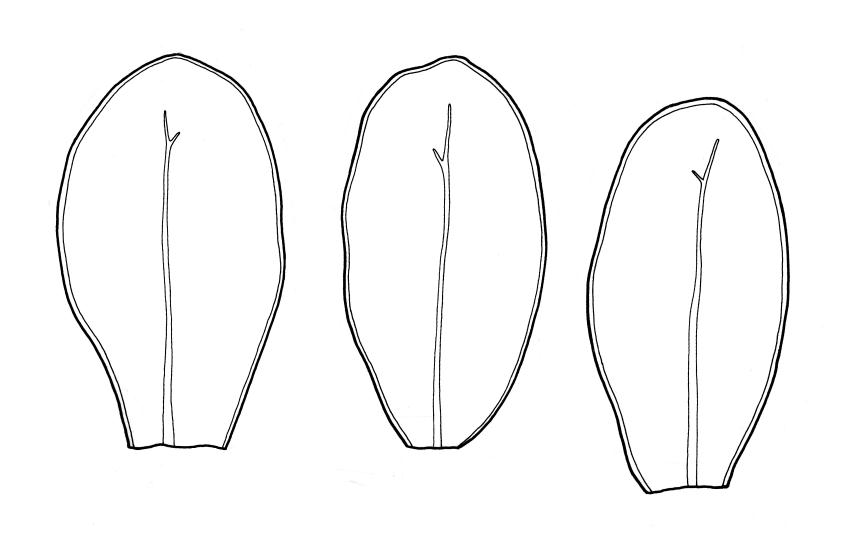
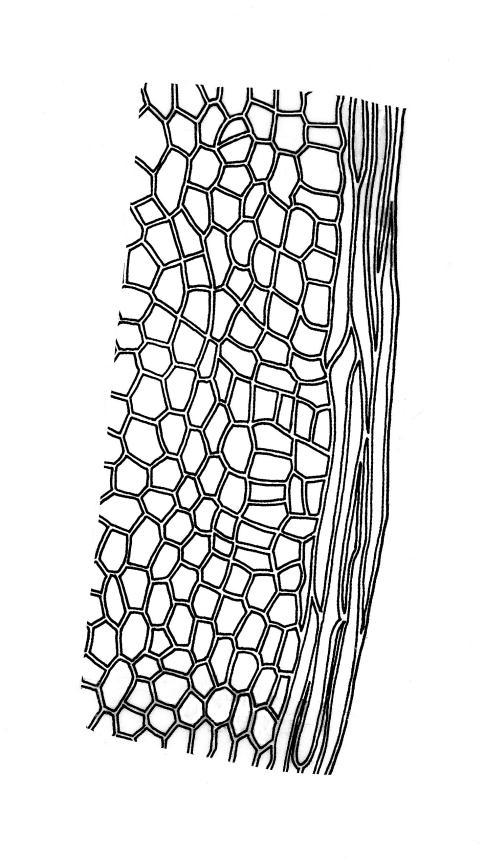
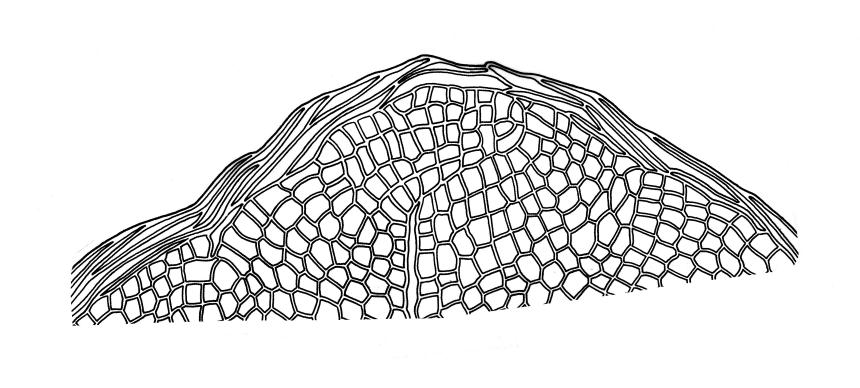
Plants medium-sized to robust, soft, yellow-green to red-brown to occasionally nearly black, variably iridescent when dry, forming dense and usually layered terrestrial or aquatic mats. Stems ± prostrate, sparsely to densely and irregularly branched, red-brown in older portions, in cross-section as per genus, c. 5–60 mm. Shoots mostly complanate, sometimes (in aquatic forms) terete, c. 2 mm wide. Leaves either inserted in c. 6 rows and complanate or neither conspicuously ranked nor complanate, symmetric or weakly asymmetric at insertion, mostly strongly crisped when dry (sometimes weakly crisped), usually obscuring the complanate nature of the shoots, variable in outline (broadly obovate, spathulate, lingulate, oblong-elliptic, or occasionally ovate), broadly rounded at apex, often with a small apiculus, weakly narrowed towards insertion, clearly or scarcely concave, plane or reflexed at margins, bordered by elongate concolorous or pigmented cells, entire, (1.0–)1.2–2.0(–2.8) × 0.7–1.0(–1.2) mm; upper laminal cells hexagonal (sometimes oblong or ± irregular), firm-walled, (6–)9–18(–24) × (5–)7–16 µm, becoming larger and more regularly hexagonal in mid leaf and larger still (to c. 60 µm) and lax towards insertion; juxtacostal cells not differentiated; border strongly defined, either 2–5 cells and 15–24 µm wide at mid leaf in terrestrial forms or 27–36 µm at mid leaf in aquatic forms, extending entire leaf length, either concolourous (in terrestrial forms) or red-brown to dark cerise (in aquatic forms). Costa concolorous and (9–)12–21(–24) µm wide in lowest ⅓ (in most terrestrial forms) or red-brown to dark cerise and 21–39 µm wide in lowest ⅓ (in aquatic forms), simple or with a lateral spur, reaching ⅔–⅞ the leaf length, usually ending 225–400 µm from the apex (or <225 µm in aquatic forms). Axillary gemmae rare, filamentous, 150–225 µm and c. 6 cells long, in dendroid clusters arising from rhizoids on the abaxial surface of the leaf base.
Dioicous. Perichaetial leaves ovate-oblong to elliptic, broadly obtuse or rounded apically, often irregularly toothed, variably (usually weakly) costate and bordered, c. 0.5–1 mm. Perigonia rarely seen, scattered on ♂ stems, c. 0.8 mm long, with ovate, obtuse, ± apiculate bracts, with several (c. 8) antheridia and lacking paraphyses. Setae (7–)10–25 mm, slender and smooth, weakly sinistrorse, red-brown; capsules horizontal or pendent, oblong-cylindric from a tapered neck c. ¼ the total capsule length, (1.0–)1.2–1.8 mm, constricted below the mouth when dry, red-brown; exothecial cells isodiametric or short-hexagonal, strongly collenchymatous; annulus apparently lacking; operculum rostrate from a conic base, c. 0.8 mm. Exostome teeth c. 350 µm, yellow- or pale brown; endostome segments arising from a high basal membrane, ± equal the teeth in height. Calyptra mitrate-rostrate, c. 1.8 mm, smooth above, fimbriate at base. Spores 9–18 µm, green, appearing smooth.
Wilson 1854, pl. 43, fig. 3 (as Hookeria amblyophylla); Sainsbury 1955, pl. 65, fig. 1; Matteri 1975a, pl. 1 (as D. kraussei); Matteri 1975a, pl. 4 (as D. fuegianum); Fife & Matteri 1984, fig. 1–4 (mostly spores, as D. kraussei), figs 5–6 (spores); Streimann 1999, fig. 11; Malcolm & Malcolm 2003, p. 24.
NI: N Auckland, including offshore islands (TK, LB, GB), S Auckland, Gisborne, Hawke’s Bay, Taranaki, Wellington; SI: Nelson, Marlborough, Canterbury, Westland, Otago, Southland; St; Ch; A; C.
Austral. Tasmania*, mainland Australia*, Argentina (Tierra del Fuego)*, Chile (Magallanes Province)*.
This is the most widespread, common, and variable species of Distichophyllum in N.Z.; it occurs widely in virtually all forest types in N.Z., ranging from coastal forest to subalpine southern beech-dominated forests and extends into the alpine zone in protected microhabitats. It also occurs in lowland wetlands such as wet dune slacks, and Typha-dominated swamps. In forests, D. pulchellum is predominantly but not exclusively a terrestrial species occurring on a range of substrates including moist duff, on shaded rocks and outcrops, decaying logs, on tree bases and trunks. It is best developed in moist but not waterlogged sites, but also occurs in ponding areas within forests. In mesic to moist forest habitats its yellow-green and layered mats of very soft, weakly complanate shoots with bordered, spathulate, apically rounded, and loosely ranked leaves are readily recognisable. Such plants characteristically lack secondary pigmentation and have narrow costae (mostly <24 µm in lowest ⅓) and narrow leaf borders (15–24 µm wide at mid leaf). Stem length is extremely variable, ranging from c. 5 to c. 50 mm, with irregular branching. The species also occurs in waterlogged sites (e.g., lake or tarn margins), as well as in swiftly flowing rivulets and waterfall margins in subalpine and lower alpine zones. One aquatic collection from a depth of 25 m in either Lake Rotoroa or Lake Rotoiti (Nelson L.D.) has been seen. It occurs in plantations of exotic trees as well as in native forests. Distichophyllum pulchellum assumes characteristic growth forms in some of these habitats (as discussed in greater detail below), some of which have been taxonomically described.
On the North I. D. pulchellum s.l. ranges from near sea level (near Wairoa, Hawke’s Bay L.D.) to greater than 1200 m elevation (Mt Taranaki and Mt Ruapehu) and on the South I. from near sea level (Measly Beach, Otago L.D.) to at least 1580 m (Mt Arthur, Nelson L.D.).
Distichophyllum pulchellum is extremely catholic in its habitat preferences and correspondingly variable in its morphology. Several infra-specific taxa have been proposed, but none are accepted here as having taxonomic value. The South American name, D. kraussei, which has been widely applied in N.Z., is likewise regarded as unworthy of taxonomic segregation. The great range of morphological variation of D. pulchellum correlates with environmental factors, particularly water availability, flow rates, and light intensity.
When D. pulchellum occurs in insolated and waterlogged habitats, it often assumes a characteristic growth form, with the shoots terete (rather than loosely complanate) and leaves oblong-elliptic (rather than ± spathulate) and not in clear ranks on the stem. The borders (at mid leaf) in such material are 18–30 µm wide, the costae are 21–30(–36) µm wide below and all these structures lack (or have minimal) secondary pigmentation. This growth form has been described from Southland L.D. as var. ellipticifolium Sainsbury (1952) and occurs widely throughout the main islands from sites near sea level to in excess of 1300 m elevation. Plants growing in non-flowing water usually do not develop strong secondary pigmentation although exceptions do occur (e.g. G.B. Huang 179 from the margin of Lake Cobb, Nelson L.D., CHR 462705). The features that distinguish it from more representative D. pulchellum are taxonomically trivial and a range of intermediates occur. The variety ellipticifolium is not recognised but is considered as an environmentally induced growth form.
Sainsbury (1947) applied the varietal name parvirete to plants of D. pulchellum with smaller-than-normal upper laminal cells. The type specimen of the variety (from South Auckland L.D.) has upper laminal cells of 6–11 × 5–9 µm, with considerable variation between leaves. These dimensions lie near the lower end of a continuous range of variation for D. pulchellum. These smaller leaf cell dimensions occur in specimens from throughout N.Z. and are correlated with thicker cell walls and strongly crisped leaves. The continuous nature of the variability precludes recognition of the var. parvirete Sainsbury. H.N. Dixon was apparently of the same opinion when he wrote to Sainsbury (4 Jan. 1935) stating that the type specimen of the var. parvirete was merely an "extremely small" celled form of D. pulchellum.
Distichophyllum pulchellum also occurs on submerged rocks or gravels in or at the margins of swift subalpine streams and in adjacent flushes. Plants from such habitats develop broader leaf margins and costae, usually with strong secondary pigmentation. The secondary pigments often suffuse the walls of the laminal cells rendering the entire plants cerise to nearly black. The individual shoots are terete, with the plants usually forming ± pendent, layered mats. The leaf borders are usually 27–36 µm wide at mid leaf and the costae are mostly >30 µm wide in their lowest third, but with extreme values overlapping with terrestrial plants. The costae in such plants tend to end closer to the leaf apex than those in representative terrestrial plants. Stoutly bordered and highly pigmented plants from high energy aquatic habitats often have eroded leaves on the lower shoots, sometimes with only the costae persisting. When viewed in isolation such aquatic material is distinctive and has been treated in both N.Z. and South America (Fife & Matteri 1984; Matteri 1975a) as D. kraussei (Lorentz) Mitt. Streimann (1999), studying Tasmanian material, rejected the value of the distinguishing features used by Fife & Matteri (1984) to segregate D. kraussei. New Zealand plants exhibiting these features occur largely near to or west of the Main Divide on the South I., to elevations of c. 1400 m. Examination of large numbers of specimens (particularly those made since the 1980s) from a wide elevational range on all three main islands shows that intermediates occur for all these features, and the distinction between terrestrial forms of D. pulchellum and the alleged N.Z. D. kraussei has become increasingly blurred. Accordingly, and following examination of South American material including the type, D. kraussei is considered to be a growth form and a taxonomic synonym of D. pulchellum. The absence of records of D. pulchellum s.s. from South America then invites explanation. Isotype material (CHR 414362) of D. fuegianum Matteri (1975b) is in my opinion referable to D. pulchellum, notwithstanding her statement in the protologue that the former is monoicous. However, further investigation of this question is beyond the scope of this Flora.
Dendroid clusters of axillary filamentous gemmae can very rarely be found in D. pulchellum, and these are similar to those Sainsbury (1952) described for the so-called var. ellipticifolium.
Mniadelphus pulchellus Hampe in Müll.Hal. is the earliest legitimate name for the present species, but Müller (1850–1851, p. 23) cited Hookeria pulchella Hook.f. & Wilson (1844) as an earlier published name. The 1844 name, however, is an illegitimate homonym preceded by the Indian Hookeria pulchella Griff.
Distichophyllum zuernii Schlieph. in Müll.Hal. (1902) and Hookeria microclada Colenso (1886) were both placed in the synonymy of D. amblyophyllum by Dixon (1927, p. 283), who saw type material of the former, but not the latter. Types of these names have not been seen for this treatment, but little purpose would be served by questioning Dixon’s synonymies.
Dixon (1927) took a narrower view of D. pulchellum than the one proposed here. He differentiated D. amblyophyllum from D. pulchellum on the basis that the former had leaves broader above, and leaf margins generally reflexed, quite obtuse or rarely with a minute apiculus. Dixon allowed for insufficient variability within the protean D. pulchellum.