Class
Family
Genus
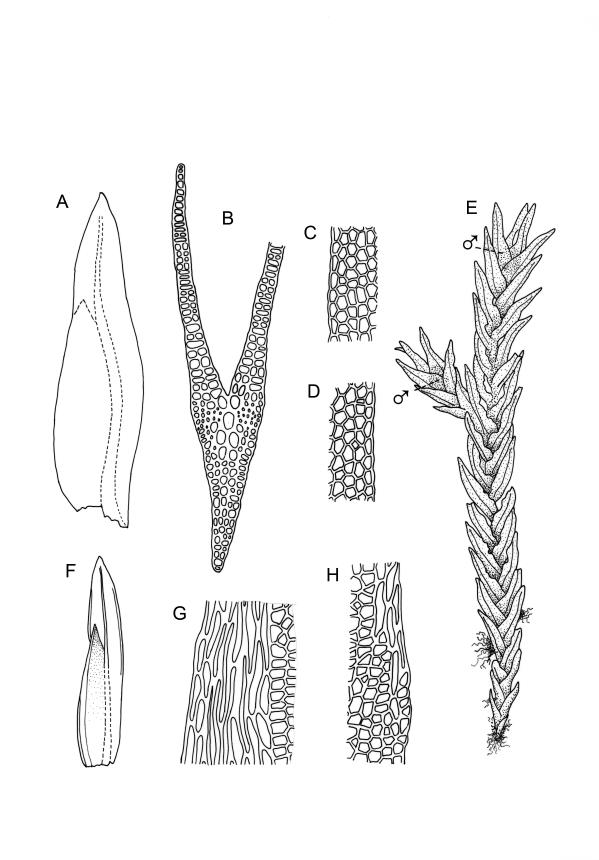
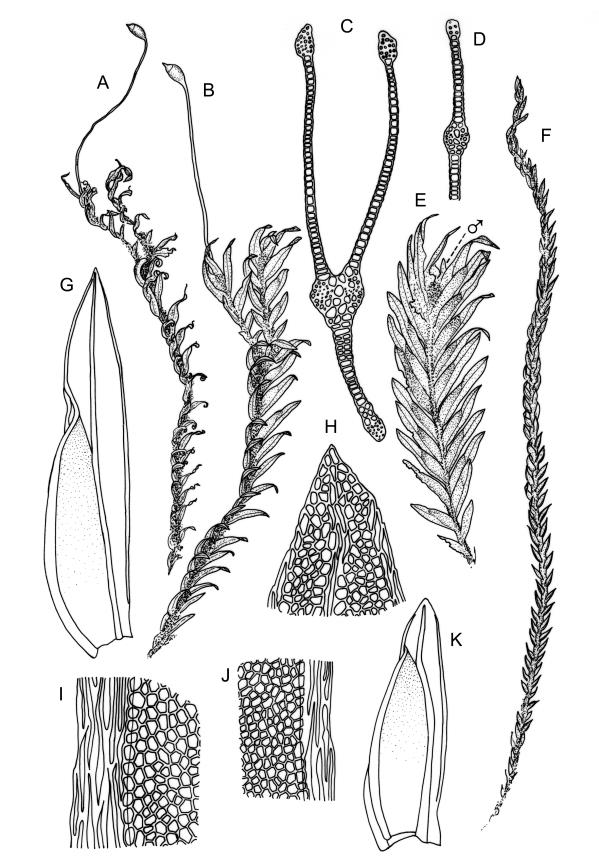
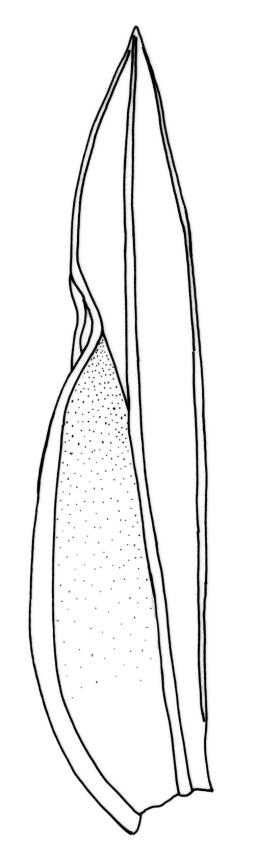
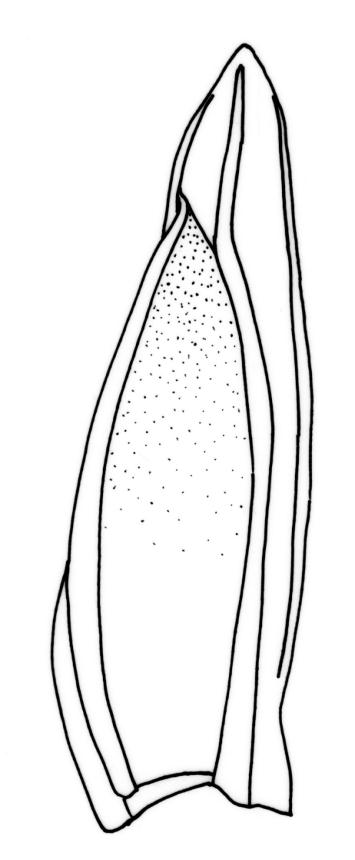
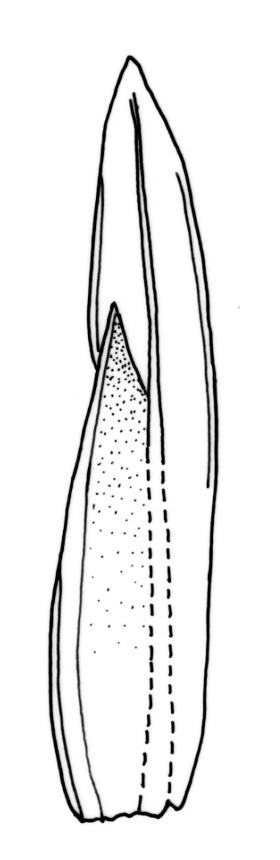
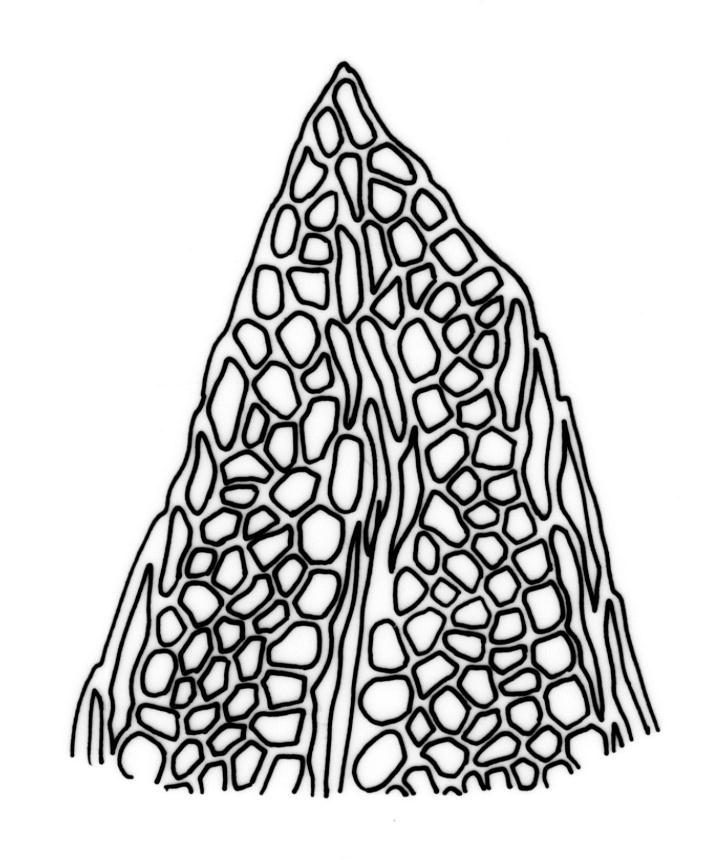
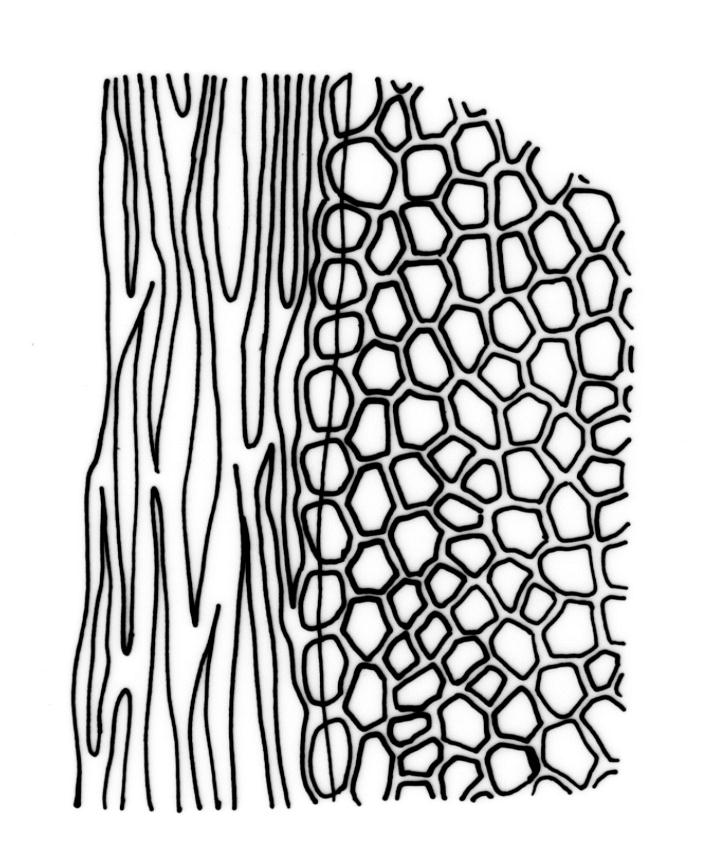
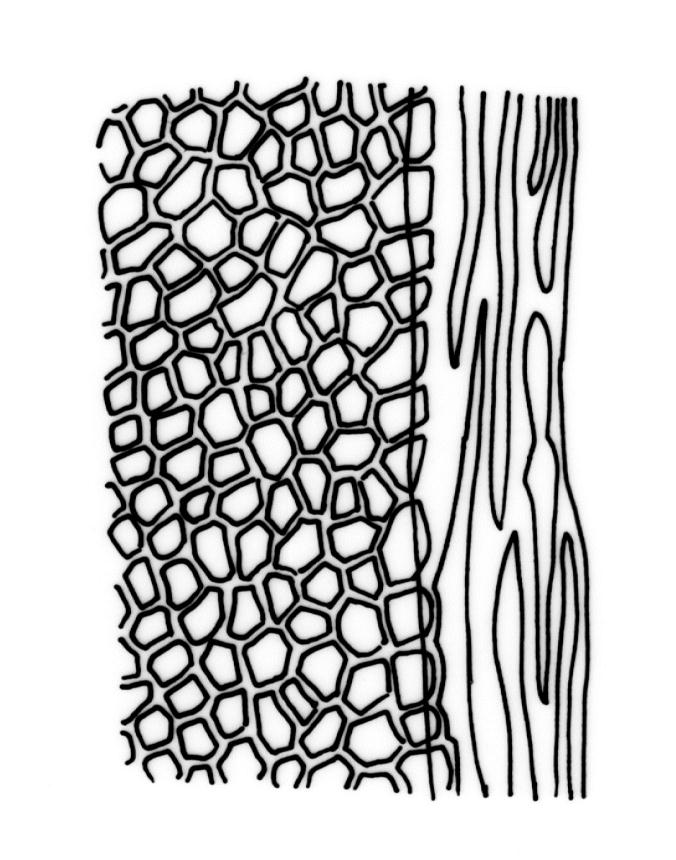
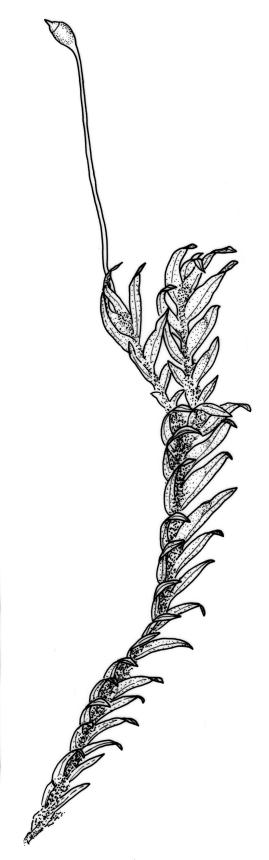
Plants robust, 10–80(–120) mm, tufted or forming dense mats. Leaves in 10–80 pairs, slightly overlapping to distant at mid stem, spreading, decurved when moist, crispate or regularly spirally twisted with apices visible (or rarely little altered, in strongly pluristratose forms) when dry, lanceolate to broadly lanceolate, 1.5–3.5 × 0.3–0.8 mm; laminae unistratose, or partially and variably pluristratose adjacent to the costa and in patches in dorsal and apical laminae; dorsal lamina reaching to leaf insertion and shortly decurrent, tapered to its base; margins entire; marginal cells on all laminae distinct in several rows, thick-walled, prosenchymatous, forming a pluristratose border on all margins except at leaf apex and base of dorsal lamina, or occasionally with border rudimentary; cells of apical and dorsal laminae (5.0–)7.5–10.5(–15.0) × (6.0–)7.5–10.5(–12.0) µm.
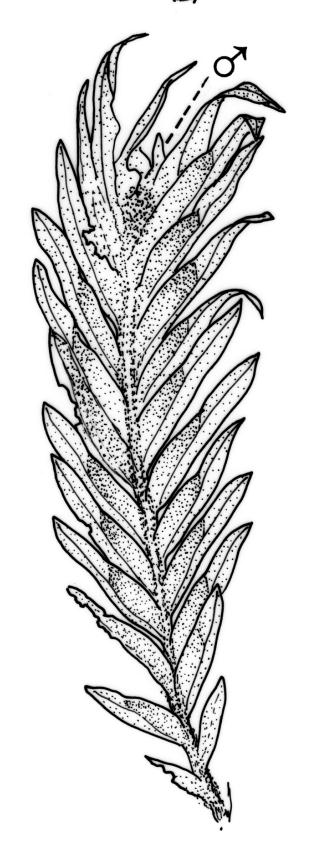
Dioicous. Perichaetia and perigonia terminal on similar-sized plants. Perichaetial leaves longer than vegetative. Setae orange-brown, wiry, 4–7 mm; capsules inclined, slightly asymmetric, 0.7–1.0 mm; operculum rostrate from a conic base, ½–⅔ the length of theca. Peristome bryoides-type; teeth with long-columnar papillae on the adaxial trabeculae below the bifurcation, 42–72 µm wide at base. Calyptra smooth, cucullate. Spores 17–20 µm.
Wilson 1854, pl. 83, fig. 3; Bruggeman-Nannenga 1974, fig. 2a; Scott & Stone 1976, pls 7–9 (pro parte); Catcheside 1980, fig. 14; Beever & Stone 1992, figs 13, 14, m; Beever & Stone 1999, fig. 2, h, n, p (as F. rigidulus var. pseudostrictus); Beever et al. 2002, p. 58, figs 1–4; Seppelt 2004, fig. 62; Stone & Catcheside 2012.
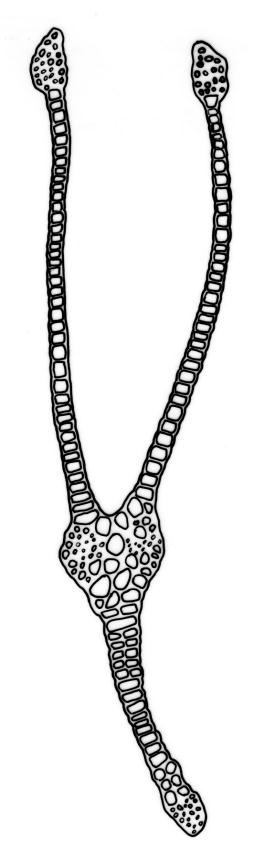
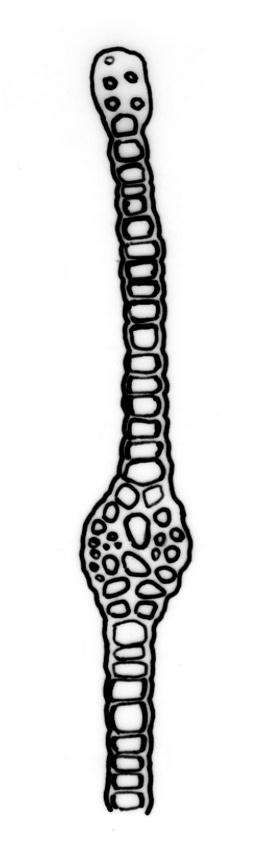
Typically the species has stout borders on all laminae, except at the extreme leaf apex and at the extreme base of the dorsal lamina. This morphological feature is no doubt responsible for a characteristic corkscrew-like leaf stance, often seen in dry shoots of F. rigidulus var. rigidulus. The stout leaf borders, constrained at either end, spiral as they contract during dehydration, and the relatively thin laminae exert little resistance to the twisting process.
Forms occur, however, in N Auckland L.D. (e.g., R.E. Beever s.n., AK 348756, Waitakere Ranges; P.J. Brownsey s.n., WELT M007840, Puketī Forest; P.J. Brownsey s.n., WELT M007797, Ōmahuta Forest) in which the vaginant laminae may have stout borders, and the dorsal and apical laminae have borders that are weak and of limited extent, or altogether absent. In extreme forms even the vaginant laminae are very weakly bordered, and no borders are seen elsewhere. Formerly these partially bordered forms were regarded as variants of F. rigidulus var. pseudostrictus (Beever & Stone 1999; Beever et al. 2002), but they are better placed, albeit mainly for convenience, in the type variety.
Another variation of leaf structure is found in F. rigidulus specimens from Te Waikoropupū Springs (e.g., D. Glenny s.n., AK 291823, Image: Image\1FQL F, K). These freshwater springs (which exhibit extreme water purity, visual clarity, and thermal stability) harbour unusual forms of several moss species (Beever & Fife 2008). Fissidens rigidulus from Te Waikoropupū Springs (Image: Image\1FQL K) has leaves with a proportionally short apical lamina, exceptionally stout leaf borders (four cells thick), and with pluristratose areas in all laminae. The resultant stiffened structure lacks the characteristic corkscrewing of leaves on drying.
Discrimination between small forms of F. rigidulus var. rigidulus and F. leptocladus is discussed under the latter species.
Bruggeman-Nannenga (1978, 1979), in studies of F. rigidulus throughout its range, commented on the great variability of the species, and noted exceptional forms with incomplete borders. Similarly, Pursell (1994) noted that in the neotropics "the species [Fissidens rigidulus] exhibits a considerable degree of morphological plasticity".
In its usual form, however, F. rigidulus var. rigidulus is the most stoutly bordered of all N.Z. aquatic Fissidens, and, with the exception of F. berteroi, the largest.
K; NI: N Auckland, including offshore islands (HC, LB, GB), S Auckland, Gisborne, Hawke’s Bay, Taranaki, Wellington (including KA); SI: Nelson (including D’U), Marlborough, Canterbury, Westland, Otago, Southland; St; Ch; A; C. Reported from M (Seppelt 2004).
Austral. Tasmania*, mainland Australia*, Chile*. Reported from New Caledonia by Müller et al. (2003), and from Argentina, Bolivia, Brazil, Falkland Is and Peru by Bruggeman-Nannenga (1979).
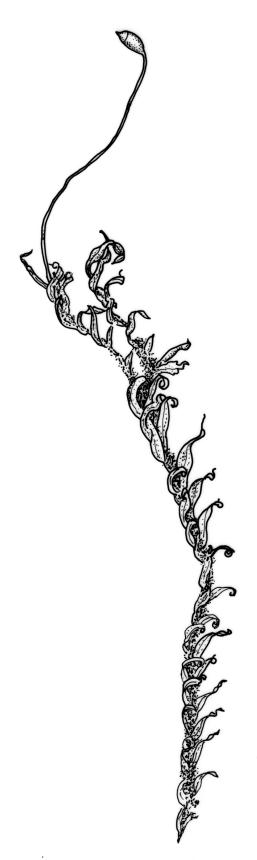
A very common aquatic moss, F. rigidulus var. rigidulus is found epilithic on a wide range of substrates, including basalt, granite, greywacke, ignimbrite, limestone, and schist, as well as on concrete. Most commonly it is a rheophyte, growing submerged in fast-flowing oligotrophic streams. It also occurs in seeps, in the splash zones of waterfalls, in swamps, and in freshwater lakes. In a detailed study of the submerged and continually wet bryophytes in 118 streams in the South I., Suren (1996) found F. rigidulus to be the most commonly collected taxon, being present at 57 sites. In a survey of deep-water bryophytes in N.Z. lakes (i.e., of those bryophytes occurring at ≥10 m depth) F. rigidulus was found to be the most frequent of over 30 bryophyte taxa recorded, occurring in 9 of the 16 lakes surveyed (de Winton & Beever 2004). It was collected in its typical form by divers from below 30 m in Lakes Sumner, Lochnager, Waikareiti, and Wanaka, and by grapnel at 50 m depth in Lake Coleridge. In addition, collections from Lake Wakatipu, from a depth range of 11–57 m, included fine green filaments, which proved to be moss stems bearing vestigial leaves; after growing on, under laboratory conditions, some were identifiable as Fissidens with borders discernible on the leaves (e.g., AK 286941), and identified as an extreme growth form of F. rigidulus.
Fissidens rigidulus var. rigidulus is tolerant of very low light intensities, such as those recorded at the bottom of deep lakes, where light levels were calculated to be 0.2–1% of incident irradiance at the lake surface (de Winton & Beever 2004), or in streams in deeply shaded gullies in forest. This moss is also found in high light conditions, as in broad forest streams under a wide canopy gap, and in exposed streams in alpine herbfields.
While most often recorded in indigenous vegetation, it can also be found in suitable sites in exotic plantations, and in urban settings. Some of these are undoubtedly relic colonies, while others, in artificial waterways in cities, must be of recent origin.
A wide range of aquatic bryophytes are associated with F. rigidulus var. rigidulus, including Achrophyllum dentatum, Aneura sp., Ochiobryum blandum, Cratoneuropsis relaxa, Echinodium hispidum, F. rigidulus var. pseudostrictus, F. waiensis, Monoclea forsteri, Philonotis australis, Platyhypnidium austrinum, Thamnobryum pandum, and Tridontium tasmanicum.
This is perhaps the only species of Fissidens in N.Z. to occur frequently above the tree-line. Records range from near sea level, even in the Far North, to over 1500 m (St Arnaud Range, Nelson L.D.).