Class
Order
Family
Aquatic (NZ) or occasionally terrestrial (not NZ). Plants growing from a lobed (in transverse section) and erect (NZ) or elongate (not NZ) corm-like stem that gives rise to a tuft of roots and a cluster of erect leaves, completely or partially submerged or occasionally terrestrial; leaves evergreen when submerged, or losing their leaves in dry periods when terrestrial. Scales absent. Leaves spirally arranged, ligulate, undivided, with a single unbranched vein surrounded by four longitudinal air canals divided by transverse septae; all leaves similar, linear, either terete and winged at base, with a subulate or acuminate apex (NZ), or flattened with a spathulate or rounded, apiculate apex (not NZ); leaf margins entire, bases swollen and colourless. Sporophylls similar to leaves, bearing sporangia that are embedded on the adaxial face of the swollen leaf base. Sporangia ovoid or globose, indehiscent, partially or completely covered by a thin membrane (velum). Heterosporous; mega- and microspores usually produced in separate sporangia (NZ) or rarely in the same one (not NZ). Megaspores trilete, with pronounced laesurae and equatorial ridge, variously patterned from almost smooth to verrucate or tuberculate, 250–800 μm in diameter. Microspores monolete, variously patterned but usually echinate, 20–40 μm in diameter.
A genus of from 192 (Troia et al. 2016) to about 250 species (PPG 1 2016).
Morphologically Isoetes is a very distinct genus and not closely related to any other lycophyte group. However, species in the genus are difficult to identify, partly because of their morphological simplicity resulting from a predominantly aquatic habitat, but also because of extensive polyploidy and reticulate evolution. Furthermore, genetic variation is limited among species of Isoetes, despite their worldwide distribution and ancient origin (Larsén & Rydin 2016).
Based on analysis of sequence data from three nuclear and plastid gene regions in 74 species, Larsén & Rydin (2016) found five major clades within the genus but struggled to correlate these with biogeographic patterns. The earliest diverging lineage (Clade A) contained species from South Africa, India, Australia and South America, and appeared to be related to ancient vicariance events, but later diverging clades appeared to be more readily explained by dispersal processes. However, the patterns were complex and require further study. Samples ascribed to the two species that have been recognised in New Zealand, I. alpina and I. kirkii, both occurred in Clade D, one of the later diverging lineages. Two different collections of each taxon were sampled, and all four showed differences; the different collections were located in separate lineages, closely related to the Australian species I. australis, I. caroli, I. humilior, and I. muelleri. This suggests that New Zealand Isoetes is genetically variable, but the available evidence does not indicate any easy way to distinguish the two currently recognised species.
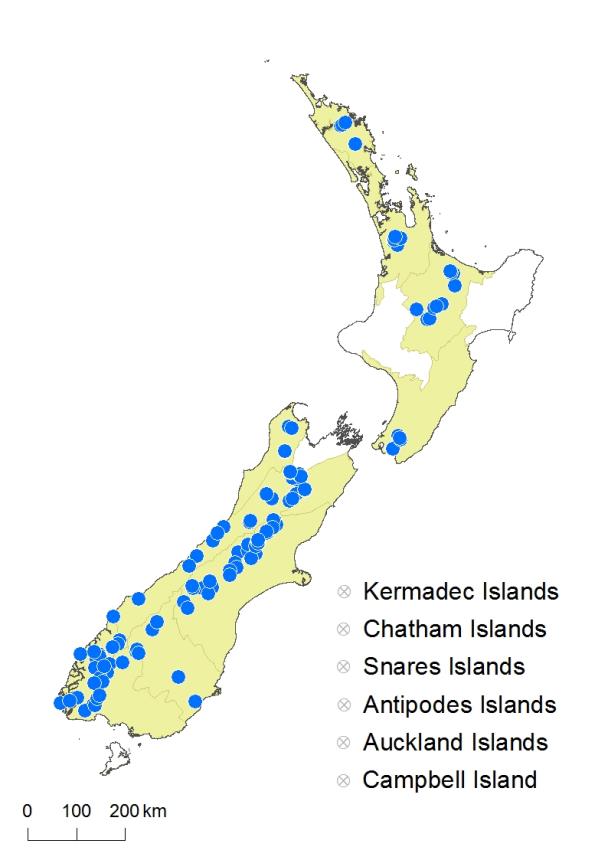
The taxonomy of New Zealand Isoetes has remained unchanged since the 19th century. Isoetes kirkii was described by Braun (1869), I. alpina by Kirk (1875) and I. multiangularis by Colenso (1890). No other names or combinations have been validly published since then. Cheeseman (1906) quickly reduced Colenso’s I. multiangularis to synonymy with I. alpina, albeit with a question mark, and the concept of two species in New Zealand was subsequently accepted by Cheeseman (1925), Allan (1961) and Brownsey & Smith-Dodsworth (2000). The two species were distinguished primarily by the megaspores, said to be smooth in I. alpina but tubercled in I. kirkii, and both species were reported from both the North and South Islands.
Since 1979 there have been several investigations of Isoetes in New Zealand, but these have either never been published, or the results have mostly appeared in conference papers, or in technical or client reports, with limited accessibility. Craig Marsden (1979) undertook a PhD thesis that investigated the morphology and taxonomy of the genus in Asia and Australasia. With respect to the New Zealand species, he proposed recognising three very similar varieties: I. kirkii var. kirkii, I. kirkii var. alpina, and I. kirkii var. flabellata. He noted that var. alpina often differed from the others in having thicker leaves, smooth megaspores, and microspores with blunt spines, but that many intermediates existed. He observed that var. flabellata was most similar to var. kirkii and differed only in having imbricate leaves arranged in a distinct flabellate pattern, fewer corm lobes (2–3 cf. 3–4 in var. kirkii), and in minor details of the megaspore and microspore ornamentation. Var. flabellata was known only from L. Ōmāpere in Northland, but he cited specimens of the other two taxa from both main islands. However, his varietal names were never published and have no standing.
In 1998 D. Brunton and D.M. Britton investigated New Zealand Isoetes, studying herbarium material, spore morphology and cytology. Their results were inconclusive and never published, but were summarised as a personal communication in a Department of Conservation Technical Report (see Hofstra et al. 2006). Brunton & Britton suggested that New Zealand Isoetes was best viewed as a single species with three similar subspecies, roughly corresponding to Marsden’s varieties. Subsp. kirkii was considered to be sexual, diploid, and largely confined to lowland areas of the North Island. Subsp. alpina was also sexual and diploid, but confined to the South Island. However, they noted that some entities on both islands were difficult to assign. They also reported (without supporting evidence) that some populations in the central North Island were tetraploid and apparently apomictic, with spores similar to, but larger than, those of subsp. alpina from the South Island.
Hofstra et al. (2006) carried out a preliminary genetic assessment of Isoetes in New Zealand for the Department of Conservation. They sampled I. kirkii from three North Island sites (including L. Ōmāpere) and I. alpina from three South Island lakes. Using ITS sequences they found little difference between any of the samples, and with rbcL sequences they reported only one base pair difference between I. alpina and I. kirkii samples. RAPDs yielded better evidence of genetically distinct entities, with the three I. alpina populations clustering together, distinct from both L. Taupō and L. Ōmāpere populations of I. kirkii. However, they concluded that further investigation was needed before any final conclusions were reached.
Hofstra & de Winton (2013, not publicly available; see Hofstra & de Winton 2016) used molecular markers to show that there were distinct differences between populations of I. kirkii from L. Ōmāpere and those in the central North Island, and between I. kirkii and I. alpina. However, they noted that these genetic differences did not provide any clarity on the taxonomy of Isoetes in the absence of clear morphological differences.
Hofstra & de Winton (2016) conducted a much larger study using RAPD profiles of 254 plants from 20 lakes to determine the level of genetic variation in New Zealand Isoetes. They showed that over 80% of the variation was partitioned between, rather than within, populations. Four clusters were recovered from UPGMA analysis; two clusters included only plants from 13 South Island lakes, one included only plants from L. Ōmāpere, and the fourth included plants from four North Island and two lowland South Island lakes. Morphological support for this latter relationship is not known, and populations of other lowland West Coast lakes clustered with other South Island populations. Populations in the northern South Island were not sampled. Hofstra & de Winton (2016) noted that there were low levels of genetic variation in L. Rotoiti, near Rotorua, and L. Taupō (compared to L. Te Anau and L. Wānaka), suggesting that this may be an indication of apomictic reproduction, as previously inferred by Brunton and Britton (see Hofstra et al. 2006) at these localities. Hofstra & de Winton (2016) concluded that the recognition of genetically distinct populations has significant implications for the conservation of New Zealand Isoetes, but offered no new taxonomic opinion.
The conclusion from investigations spanning 40 years is that although there is evidence of genetic variation between populations of Isoetes across New Zealand, the variation has not yet been correlated with morphological differences. In particular, it is not clear whether there is a clear boundary between I. kirkii and I. alpina or where it may lie. The most genetically distinct population appears to be that from L. Ōmāpere (now believed to be extinct in the wild, and known only from material in cultivation), but morphologically this seems less easy to distinguish from I. kirkii than is I. alpina. There are also suggestions (Hofstra et al. 2006) that some populations from L. Taupō and L. Rotoiti are tetraploid, lack microspores and are apparently apomictic, although the evidence for that has not been published. Colenso’s I. multiangularis came from L. Taupō, so if the tetraploid apomicts were shown to be a different species, a name might be available.
The genetic evidence is, at best, inconclusive; there could be one variable species or perhaps three or four different entities. However, because these entities have not yet been correlated with morphological differences, it is unclear whether they warrant taxonomic recognition, and it would be premature to give them any formal rank. Their relationship to taxa in Australia is also unresolved. We therefore retain the status quo here and accept the two species, I. alpina and I. kirkii, that have been recognised in New Zealand since the late 19th century. If subsequent work shows that more than one entity can be recognised in New Zealand, three published names are available at specific rank, or alternatively the way is clear to publish new taxa at subspecific rank without any confusion from pre-existing concepts.
Isoetes is a genus of up to 250 species (PPG 1 2016), mostly occurring in damper habitats such as marshy soil and swamps, springs, seepage channels, seasonal pools, flowing water, alpine tarns and permanent lakes, from sea level to over 4000 m (Jermy 1990; Chinnock 1998); 16 species in southern South America (Zuloaga et al. 2008), 13 in southern Africa (Crouch et al. 2011), five in China (Zhang & Taylor 2013), 15 in Australia (Chinnock 1998), and one in Hawai‘i (Palmer 2003), but absent from the other Pacific islands. Two endemic species in New Zealand.
| Category | Number |
|---|---|
| Indigenous (Endemic) | 2 |
| Total | 2 |
The base chromosome number in Isoetes is x = 10 or 11, with many polyploids recorded (Jermy 1990).