Class
Family
Genus
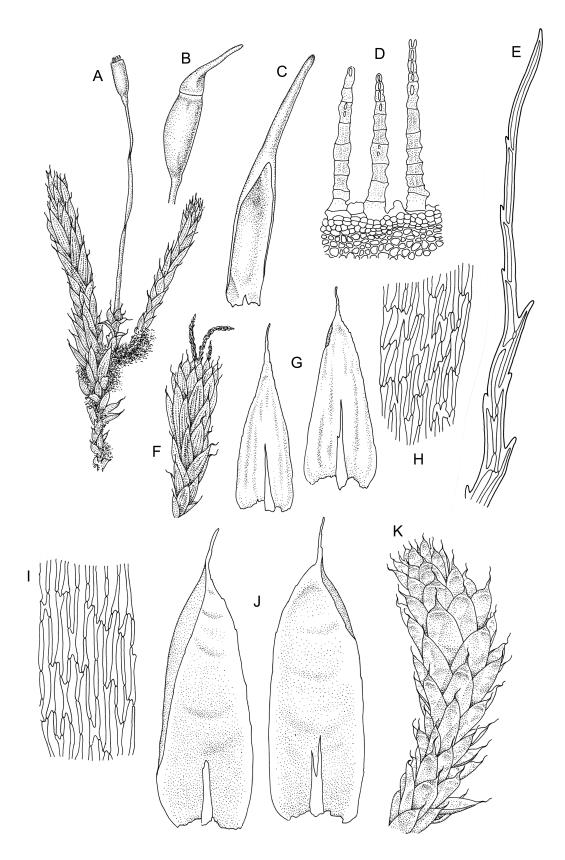
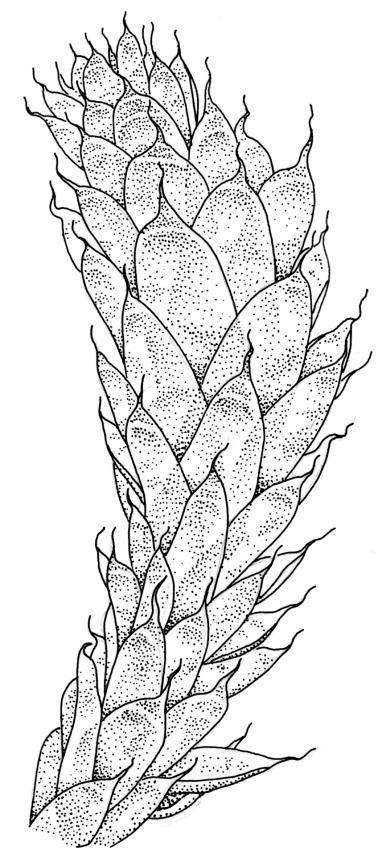
Plants soft, yellow- or brown-green, not glossy, forming extensive cushions to 0.5 m or more in diameter, terrestrial or epiphytic. Stems creeping and much branched, forming a loosely intertwined mat and giving rise to numerous erect branches, red-brown, in cross section with 2–3 layers of thick-walled cortical cells and a weak central strand, densely beset and bound together by rust-coloured, smooth rhizoids. Branches ± erect, sparsely branched, to c. 20 mm (rarely to 50 mm or more); microphyllous branches with small (c. 0.5–1.0 mm), non-plicate, ecostate leaves occasionally present, usually sparse and fragile, sometimes abundant. Leaves not ranked, imbricate, erect-spreading, symmetric, not plicate, rugose in upper half (often with convex areas at back of dry leaf), broadly oblong or oblong-lanceolate, abruptly tapered to a slender acumen up to ⅓ the total leaf length, strongly concave, serrulate in upper half or more, (2.0–)3.0–3.5(–4.0) × 0.9–1.4 mm (without acumen; reduced in size on lower portion of branch); acumen c. 0.5–1.0 mm; mid laminal cells firm-walled, vermicular, porose, mostly 54–90 × 6–9 µm, somewhat longer in lower portion of leaf, but not otherwise differentiated; cells at insertion shorter and wider; alar cells not differentiated. Costa variable, stout or thin, often bifurcate or spurred, extending ¼ to ½ the length of the lamina. Axillary hairs difficult to observe, apparently 3–4 cells long in N.Z. material.
Apparently dioicous. Perichaetia scattered on lower branches, c. 1 mm long, but elongating to c. 3 mm following fertilisation, with leaves broadly ovate-acuminate, tubular, ecostate and spreading at apices. Male plants not observed. Setae 13–23 mm, straight, not twisted, red-brown; capsules variable in length, oblong-cylindric from a short, rugose neck, (1.5–)1.8–2.8 mm, pale red-brown; exothecial cells not thickened in corners; annulus persistent, composed of a single row of inflated cells; stomata superficial; operculum as per genus, 1.0–1.5 mm. Exostome teeth rudimentary or sometimes absent; endostome with basal membrane extending 75–130 µm past the capsule mouth; segments linear, 300–520 μm long, keeled, narrowly perforate nearly throughout, not anastomosing, finely papillose-lirate throughout (but often becoming ± smooth in older material). Calyptra 4–5 mm, split to c. ½ its length. Spores 12–17 µm, smooth, pale brown.
Brotherus 1925, fig. 515, A–G; Beever et al. 1992, fig. 57; Allen 1999, figs 8–9. Seppelt has provided an informative and elegant illustration of this species in Australian Mosses Online (as L. pseudolagurus).
When dry, the rugose leaves, while strongly convex overall in abaxial view, often have a concave area in the upper third of the leaf. The leaves are markedly smaller on lower portions of the branches, and highly reduced on lower portions of main stems.
NI: S Auckland (Maungaorangi), Gisborne (near Matawai, Lake Waikaremoana), Wellington; SI: Nelson, Marlborough, Canterbury, Westland (Camp Creek, Victoria Range, Ōtira River), Otago, Southland. Vitt (1974) reported this species from C.
Austral. Tasmania*, mainland Australia*, Argentina*, Chile*. The significance of Scott & Stone’s (1976) report from "Antarctica" is unclear.
Occupying a wide range of substrates but most frequently on either sheltered or fully exposed rock (e.g., limestone, gneiss, greywacke, and granite). Also epiphytic (on Fuscospora cliffortioides and/or F. solandri, Lophozonia menziesii, Griselinia littoralis), on humic soil in rock crevices, and occasionally on rotten wood. Occurring in both forested and alpine (boulder fields, herbfields, and tussock grassland) situations. On the North I documented from scattered localities from Maungaorangi (an outlier of the Kaimanawa Range) southward, with most localities in Wellington L.D. Occurrences range in elevation from 680 m (near Matawai, Gisborne L.D.) to 1300 m (Ōhutu Ridge, Wellington L.D.). On the South I, occurring between near sea level (Tahakopa Bay Scenic Reserve, Otago L.D.) and 2000 m (Mt Tapuaenuku, Marlborough L.D.), but relatively uncommon below c. 600 m. Although found in all land districts on the South I., it is known from few localities in Westland L.D. It is, however, documented from many localities in wetter parts of Southland L.D., including Fiordland National Park. It is not documented from either Stewart or Auckland Is but has been recorded without detail from Campbell I.
Allen’s (1999) proposal to segregate N.Z. material of L. lagurus (as P. pseudolagurus B.H. Allen, Bryobrothera 5: 40) from the South American species is not accepted here, although Klazenga (2012) followed this proposal in his treatment for Australia. Allen found many gametophytic characters to be "so variable they cannot be used to separate the two", but he considered material from South America to be a "somewhat smaller plant with a more weakly developed stem central strand" than Australasian material. He also found the axillary hairs to be 3–4 celled in South American material and 4–6 celled in Australasian material and cited differences in the degree of immersion of the stomata and in the presence/absence of a rudimentary exostome. The last feature was a distinction that he chose to emphasise in his key to species. All the features he used to segregate Australasian material are either ill defined, difficult to observe, or both. The majority of collections from both regions available for examination lack sporophytes, and even in some fruiting material observations on the nature of an exostome are difficult. Most South American material indeed seems to lack an exostome (as predicted by Allen), but other collections (e.g., CHR 456548; M.R. Crosby 13057 from Prov. Malleo in southern Chile) clearly possess a rudimentary exostome and some N.Z. material also lacks an exostome (CHR 461964, B.H. Macmillan 89/17 from Lake Peel, Nelson L.D.). Thus no clear distinction can be drawn using the nature of the exostome, nor can it be correlated with geographic distribution. The nature of the stomata is likewise difficult to observe in many fruiting specimens, albeit with some clearly not conforming to Allen’s predictions. For example, A.J. Fife 7471 (CHR 106562 from Hope Range, Nelson L.D.) has clearly superficial stomata, rather than the immersed stomata predicted by Allen.
Other differences cited by Allen (1999) appear to overlap or are equally non-convincing. Axillary hairs are here extremely difficult to observe; the few observed (seen only after staining) in N.Z. material (CHR 461964) appear to be 3–4 cells long and thus conform to Allen’s description for L. lagurus rather than for the segregate species.
Male plants have not been seen. A single collection in CHR (R. Mundy 382 from Mt Ruapehu, CHR 645985) was annotated by B.H. Allen to contain males, but close examination failed to locate perigonia. Allen’s suggestion that ♂ plants are not dwarfed in this species is not accepted here. On the weight of available evidence, the continued application of the name L. lagurus to N.Z. material is desirable.