Class
Family
Genus
Tufted 15–55 mm in height. Apomictic or sexual, plants in populations often female only, or plants cosexual with unisexual or bisexual reproductive units. Leaves 14–55 × 0.25–0.4 mm. Reproductive units 1–4 per tuft, on scapes 20–40 × 0.3–0.4 mm; involucral bracts 2–4(–5) ovate to narrow-ovate. Male reproductive unit bracts 3.5–5.0 mm long; stamens (1–)3–8; anthers 0.8–1.4 mm long, bright red, filaments 1–5 mm long. Bisexual reproductive unit bracts 4–5 mm long; stamens 1–5; carpels 2–10. Female reproductive unit bracts 2.5–5.0 mm long; carpels 8–24, reddish, with 5–13 stigmatic hairs of unequal length, 0.3–1.0 mm long, red becoming hyaline. Fruits 0.4–0.56 × 0.2–0.4 mm, elliptic to ovoid.
Distinguished from T. inconspicua subsp. brevistyla by the carpels with obvious multiseptate stigmatic hairs to 1 mm long compared with a capitate head with much reduced stigmatic hairs and fruits elliptic to ovoid in shape rather than elliptic to globose.
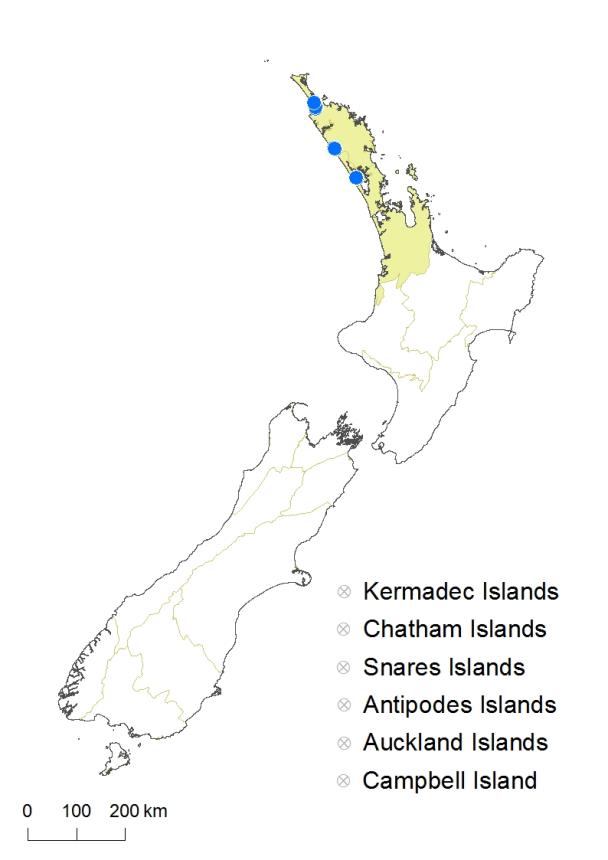
North Island: Northland – In dune lakes behind coastal dunes of the west coast between 34.9° and 36.5° latitude (Lakes Ngatu, Rotoroa, Waikare, Taharoa, Kai Iwi, Rotokawau).
In dune lakes to a depth of about 1 m (occasionally exposed above the water in a dry season, Edgar 1970); 20–70 m a.s.l. Growing in sand and silt, occasionally in peaty sediment, often part of the aquatic turf community and sometimes amongst taller sedges in the shallows.
Nationally Critical B (3/1). Reported to have declined since 1998, with lake-wide extinction in seven of the thirteen lakes reported to have supported populations (Smissen et al. 2019).
Flowering Oct.–Jan.; Fruiting from Jan. onwards.
2n = c. 24 (de Lange et al. 2004; AK 253948).