
- Taxon
- Weed
Classification
Class
Family
Genus
Subordinate Taxa
Nomenclature
Scientific Name:
Cotoneaster glaucophyllus Franch., Pl. Delavay. 3, 222 (1890)
Synonymy:
Type: China, Yunnan Province, Kiao-che-tong, near Hee-chan-men, 6 Dec. 1884, J. Delavay 3747. Holotype P 601215, image seen. Isotype: P 601216, image seen. K 758567 may be material removed from the holotype.
Description
Erect shrub 1.3–4 m tall, evergreen. Branches arched; branchlets red-brown to purple-brown, peeling cuticle present, branchlet hairs dense on young branches, becoming glabrous; stipule 5.0–6.5 mm long, green turning red, stipule hairs dense to sparse, white. Leaves sparse to crowded on branches, 30–71 mm long, lamina (32)36–69 mm long, (18)24–33 mm wide, elliptic, plane or V-shaped, sometimes slightly channelled and slightly to distinctly cupped at apex, 250–350 µm thick, petiole 3–10 mm long, sparse to dense, white; leaf base obtuse; leaf apex obtuse, apiculus straight; lateral veins in 4–9 pairs, midvein and base of lateral veins impressed above and projecting below, upper surface smooth to very slightly rugose, dark to moderate yellowish-green (RHS 139A–B), dull to glossy, upper surface hairs none or sparse; margins plane to very slightly recurved, hairs dense on young leaves, none on old leaves; lower surface glaucous, hairs dense, white, straight, fine, appressed, not persisting on old leaves.
Inflorescence a compound corymb of 16–104 flowers, 55–140 mm long, rachis hairs dense, white; pedicel 1.7–3.5 mm long, hairs dense, white. Flowers 6.5–9.0 mm diameter. Hypanthium green, sepals 1.2–1.7 mm long, 1.4–2.0 mm wide, hairs dense, straight, fine, white to pale yellow. Petals white in bud, when open spreading, white, 2.5 mm long, 2.8 mm wide, base slightly clawed, tuft of hairs usually present, margins entire or toothed. Stamens 11–20; filaments 2.1–2.8 mm long, white; anthers dark pink to red-purple, 0.5–0.9 mm long; styles invariably 2, 1.6–2.9 mm long. Fruit vivid reddish-orange (RHS 33A), oblate to obovate, 4.5–5.4 mm long, 4.5–6.4 mm diameter, calyx closed or open, glabrous. Pyrenes invariably 2, 3.3–4.3 mm long, 2.6–3.0 mm wide, hairs moderately dense at apex, umbonate, style attached at pyrene apex or up to 0.6 mm below apex.
Taxonomy
Lu & Brach (2003) recognise four varieties in Cotoneaster glaucophyllus. Var. serotinus is distinguished by having inflorescences with white tomentum and up to 40 flowers, rather than pale yellow tomentum and up to or more than 50 flowers in var. glaucophyllus. The two other varieties, var. vestitus and var. meiophyllus, have smaller leaves (15–30 mm long rather than 30–60 mm).
Fryer & Hylmö (2009) describe C. glaucophyllus as having the hypanthium glabrous except at base, while C. serotinus is described as having dense silky hairs on the hypanthium (visible on the type). All N.Z. material has dense hairs on the hypanthium and corymb branches and therefore matches C. serotinus rather than C. glaucophyllus (sensuFryer & Hylmö 2009). I have not seen any herbarium material of C. glaucophyllus var. glaucophyllus other than the type, which is in the fruiting stage. The type of var. glaucophyllus almost completely lacks hairs on the corymb branches, consistent with the description of Fryer & Hylmö (2009).
The name Cotoneaster glaucophyllus f. serotinus has been in common use in New Zealand (e.g., Given 1982).
Recognition
Evergreen erect shrub. Leaf lamina 30–60 mm long, upper surface glossy, lateral veins lightly impressed, lower surface glaucous, in young leaves densely hairy but hairs soon lost to become only sparsely hairy with fine white or silvery hairs. Flowers in compound corymbs of usually 15–40 flowers but rarely up to c. 100 flowers. Petals white, spreading, with a small hair tuft. Filaments white, anthers purple. Fruit relatively small (4.5–5.5 mm in diameter, most other species have fruit 6–10 mm in diameter), reddish-pale yellow or dull red, globose, oblate or oblong, glabrous except at base and calyx. Pyrenes invariably 2 per fruit, style at pyrene apex or up to 0.4 mm below.
Most similar to Cotoneaster coriaceus. Both are large shrubs, eventually becoming small trees, with arching branches; both have leaves with few or no hairs on the upper surface and dense tomentum on the lower surface of new leaves that disappears so that old leaves have sparse hair to none on the lower surface. Both have compound corymbs of flowers that have white spreading petals that commonly have a hair tuft. Stamens number in both species are c. 20; filaments are white and anthers red-purple. Both have 2 styles per flower and 2 pyrenes per fruit. Fruit in both species are relatively small (4–6 mm long), with pyrenes that have the style attached only slightly below the pyrene apex. Both mature their fruit in autumn (late March to April) and so are the last species to ripen their fruit.
Cotoneaster glaucophyllus differs in the lateral leaf veins, being either lightly impressed or not impressed, and the old leaves are almost glabrous on the undersurface (sparsely hairy in C. coriaceus). The leaves are often yellow-green on the upper surfaces while in C. coriaceus they are usually mid to dark green. Plants of C. glaucophyllus with mid green leaves are also common and are more difficult to distinguish from C. coriaceus, particularly in early summer when hairs are dense on the leaves of both species. The leaf and stem hairs in C. glaucophyllus are white or appear silvery, while in C. coriaceus they are pale yellow in early summer, becoming white or silvery late in summer. The leaves of C. glaucophyllus are elliptic, while in C. coriaceus they are obovate. In C. glaucophyllus they are slightly folded or channelled (plane in C. coriaceus), and the margins are not recurved (recurved in C. coriaceus).
Similar to Cotoneaster frigidus in having leaf undersides that are densely woolly-hairy in young leaves but become sparsely hairy with age, and in having many flowers and fruit per corymb. Leaves are on average smaller, lamina (32)36–69 mm long in C. glaucophyllus rather than 71–114 mm long in C. frigidus.
Distribution
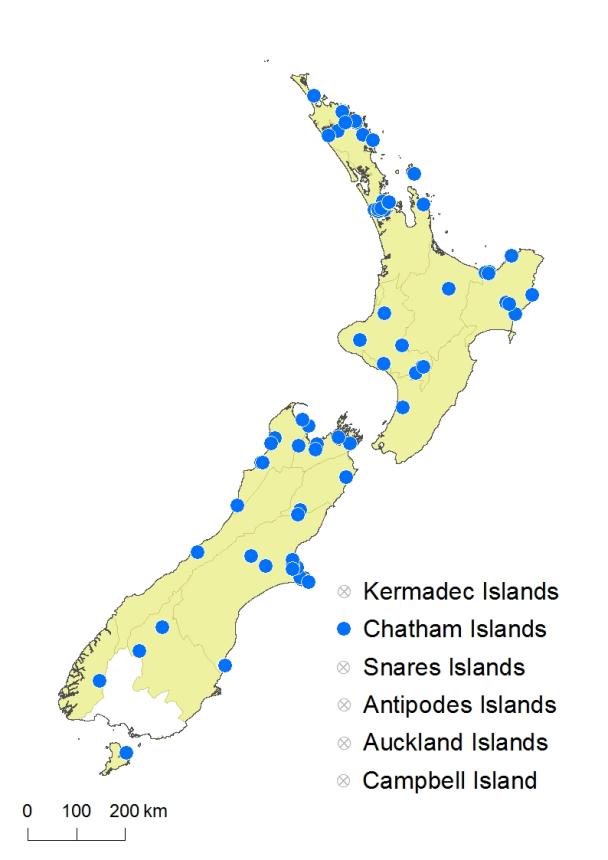
Northland (Perpendicular Point, 1992; Kaikohe, 1984; Kaio, 1975; Ōpua, 1986; Waimate North, 1981; Paihia, 1988; Whakapara, 1987; Ngunguru, 1987; Baylys Beach, 1970), Auckland (Waimamaku, 1989; Browns Bay, 1979; Campbells Bay, 1978; Auckland City, 1976, 1987; Rangitoto Island, 1994; Great Barrier Island, 1986; Titirangi Beach, 1987; Waitakere Range, 1974; Whitianga, 1985), Gisborne (Whanarua Bay, 1997; Tolaga Bay, 2001; Ōpōtiki, 1975; Ngātapa, 1989; Hangaroa, 1989), Volcanic Plateau (Mt Tarawera, Waiotapu, 1979; Raetihi, 1996), Taranaki (Ahitītī, 1983; Waitaanga Plateau, 1998; Stratford, 1977; Taihape, 1993; Mangaweka, 1997; Rangitikei R, 1993), Southern North Island (Maxwell, 1994; Kōputaroa, 1975), Western Nelson (Karamea, 1985; Ngākawau, 2008; Ngākawau Gorge, 1975, 1979; Westport, 1977), Sounds–Nelson (Nelson City, 1986; Aniseed Valley, 1991; Torrent Bay, 1996; Pelorus Sound, 1999; Picton, 1997), Marlborough (Waimā R., 2010), Westland (Dunollie, 1965; Ōkārito, 2003), Canterbury (Hanmer, 1988; Christchurch City, 1987, 1993; Port Hills, 1986, 2001; Sumner, 1987; Cooptown, 1987; Hinewai, 1995; Hororātā, 1989; Lake Coleridge, 1998), Otago (Hāwea, 2001; Queenstown, 1989; Moeraki, 1987, 2003; Waipori Falls, 1977), Fiordland (Lake Manapōuri, 1997), Stewart Island (Halfmoon Bay, 1980, 2008), Chatham Islands (Waihī, 2007; Kaingaroa, 2006; Owenga Road, 2007). Common in cultivation throughout New Zealand.
Biostatus
Exotic
| Category | Number |
|---|---|
| Exotic: Fully Naturalised | 1 |
| Total | 1 |
First Record
CHR 157684, I. Robbins, July 1965, Dunollie, in Given (1982) as C. glaucophyllus forma serotinus (Hutch.) Stapf
Phenology
Flowering: early September, peaking late November to early December, rarely late December; Fruiting: February to July, not persisting over winter
Cytology
Tetraploid, based on flow cytometry results from three plants in cultivation: CHR 638088, CHR 638100, and CHR 638089. Ploidy not stated by Fryer & Hylmö (2009).
Images







Bibliography
Biosecurity New Zealand 2012: Regional Pest Management Strategies Database. http://www.biosecurityperformance.maf.govt.nz/
Franchet, A.R. 1890: Plantae Delavayanae. Part 3. P. Klincksieck, Paris.
Fryer, J.; Hylmö, B. 2009: Cotoneasters: a comprehensive guide to shrubs for flowers, fruit, and foliage. Timber Press, Portland.
Given, D.R. 1982: Checklist of dicotyledons naturalised in New Zealand 13. Rosaceae. New Zealand Journal of Botany 20: 221–226.
Howell, C. 2008: Consolidated list of environmental weeds in New Zealand. DOC Research & Development Series 292: 42.
Lu, L.T.; Brach, A.R. 2003: Cotoneaster. In: Flora of China. Vol. 3. In: Wu, C.Y.; Raven, P.H.; Hong, D.Y. (ed.) Flora of China. Science Press, Beijing, and Missouri Botanical Garden Press, St. Louis. 85–107.