Class
Subclass
Order
Family
Rhizomes long-creeping, up to 400 mm long (in herbarium material), 3–5 mm diameter, scaly. Rhizome scales ovate, 1.5–3 mm long, 0.5–1 mm wide, brown or dark brown, margins entire or fimbriate. Fronds 240–960 mm long. Stipes 100–560 mm long, red-brown proximally, yellow-brown distally, scaly proximally, hairy throughout; scales ovate, brown or dark brown, 3–7 mm long, 1–2 mm wide, apices acute, margins entire or fimbriate; hairs whitish, catenate (ctenitoid), up to 0.6 mm long. Rachises pale brown or yellow-brown, sulcate, narrowly winged only at distal end, the sulcus filled with whitish catenate hairs up to 0.4 mm long, with slightly longer hairs up to 0.5 mm long at costa/rachis junctions. Laminae 3–4-pinnate, ovate or broadly ovate, 120–400 mm long, 90–320 mm wide, pale green on both surfaces, herbaceous; abundant whitish catenate hairs up to 0.5 mm long on both costa surfaces and on the adaxial lamina surface, more scattered on abaxial lamina surface; tiny yellow or orange clavate glands on both lamina surfaces. Primary pinnae in 14–22 pairs below pinnatifid apex, slightly overlapping; distal pinnae narrowly ovate, proximal pinnae ovate or broadly ovate; the longest at or near the base, 55–210 mm long, 30–150 mm wide, apices acuminate, bases stalked; costae narrowly winged in distal half. Acroscopic secondary pinnae decreasing evenly in length along the primary pinnae to the distal end; the basal basiscopic secondary pinna the longest, ovate, 28–115 mm long, 9–37 mm wide, apices acute, bases stalked, costae winged throughout. Tertiary segments ovate, 6–25 mm long, 2–11 mm wide, apices acute or obtuse, margins serrate to deeply divided, bases adnate or stalked. Ultimate segments narrowly oblong, up to 8 mm long and 2 mm wide, apices obtuse and toothed, margins serrate, bases adnate or stalked. Sori round, medial; indusia reniform, 0.5–1.3 mm diameter, concolorous, minutely hairy on the margins and surface.
Parapolystichum microsorum is characterised by a long-creeping rhizome, 3–4-pinnate laminae, and abundant whitish catenate hairs on both lamina surfaces. It is distinguished from P. glabellum by its creeping rather than erect rhizome, and whitish hairs on both lamina surfaces, rather than reddish hairs, which are largely confined to the adaxial surface. The abaxial rachis surface is densely hairy, unlike those in P. glabellum and P. kermadecense, which are glabrous or only sparsely hairy.
Parapolystichum microsorum in New Zealand is very similar to the plant that occurs in Australia. However, Tindale (1965) distinguished them as separate subspecies, with subsp. microsorum occurring in Australia and subsp. pentangulare (Colenso) Tindale confined to New Zealand. Subsp. pentangulare was said to have smaller laminae (140–400 mm long, 90–320 mm wide) compared to subsp. microsorum (180–500 mm long, 120–470 mm wide), with the ultimate segments more sharply spinulose-dentate (Tindale 1965). The recognition of two subspecies was described as “somewhat borderline” by Tindale (1965), but there is a difference in chromosome number, Australian plants being diploid and New Zealand plants tetraploid. The status of the species in both countries needs further investigation.
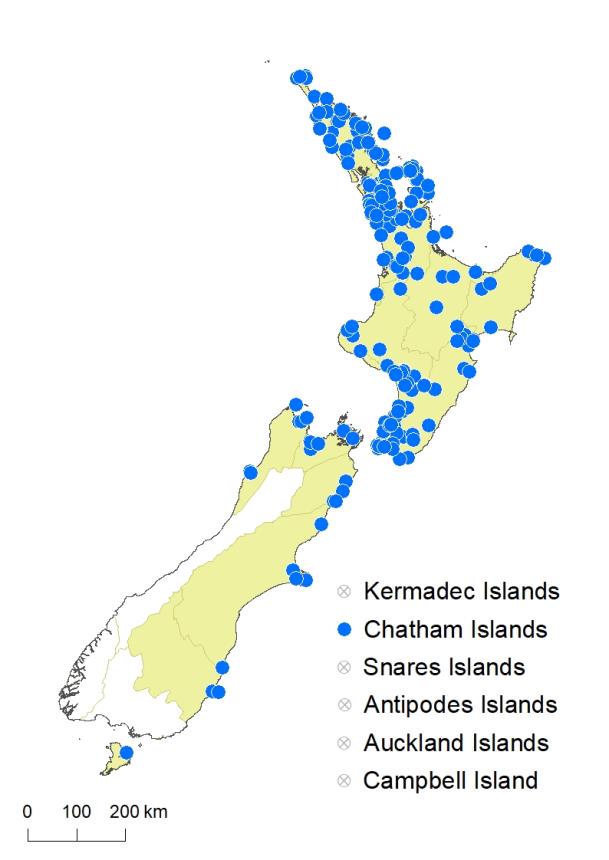
North Island: Northland, Auckland, Volcanic Plateau, Gisborne, Taranaki, Southern North Island.
South Island: Western Nelson, Sounds-Nelson, Marlborough, Canterbury, Otago.
Chatham Islands, Stewart Island.
Altitudinal range: 0–500 m.
Parapolystichum microsorum occurs in lowland areas throughout the North Island from Te Paki to Wellington, extending locally into montane areas but absent from most of the central high country. It ranges from near sea level, up to 500 m at Ngongotahā near Rotorua. In the South Island it is confined to coastal and lowland areas from Golden Bay to the Marlborough Sounds, with outlying populations south to Westport, Banks Peninsula, and Trotters Gorge near Moeraki. It reaches 150 m in the Kekerengu Valley, Marlborough, and extends to the Chatham Islands. There are early collections made by W.A. Scarfe from the Leith Valley, Dunedin (CHR 283522), and by Petrie from Otago Peninsula and Stewart Island (WELT P008540, P028836). The species was not recorded from Stewart Island by Wilson (1982), and its occurrence there, and in Otago, needs confirmation.
Also Australia (Queensland, New South Wales, Victoria).
Parapolystichum microsorum occurs in podocarp, broadleaved and coastal forest, under kānuka, and rarely under willows. It is a terrestrial fern found on the forest floor, on streambanks, on river terraces, near waterfalls, in damp gullies, amongst rocks, on clay banks, and on tracksides. It grows on greywacke, limestone and acidic volcanic substrates.
In New Zealand Parapolystichum microsorum has been reported as tetraploid with n = 82 (Brownlie 1961– as Ctenitis decomposita). In Australia it has been reported as diploid with n = 41 (Tindale & Roy 2002).
The name Nephrodium decompositum R.Br., and combinations based on it, used in earlier Flora treatments are misidentifications of Lastreopsis decomposita (R.Br.) Tindale, which is a diploid Australian species.
The name Parapolystichum microsorum subsp. pentangulare, and earlier combinations based on the same type, cannot be used for the New Zealand plant because Colenso’s original collection is a specimen of Lastreopsis velutina (Brownsey et al. 2019). Plants in Australia and New Zealand need further investigation to see if there are consistent differences between them, and whether diploid and tetraploid forms are each confined to one or other country. If New Zealand plants deserve recognition, the varietal name Nephrodium decompositum var. pubescens is available.