Class
Family
Genus
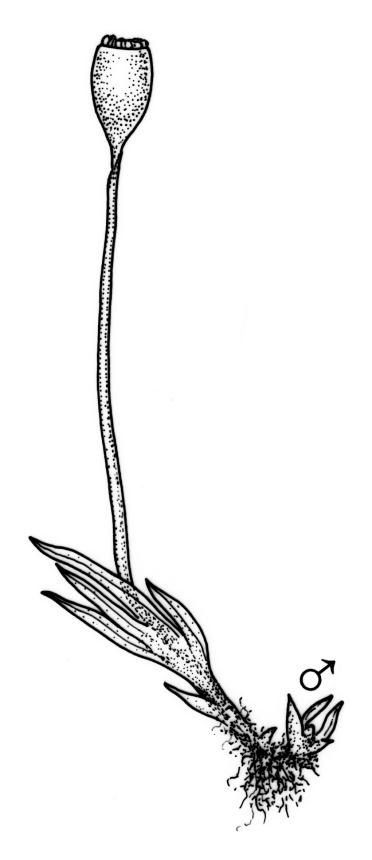
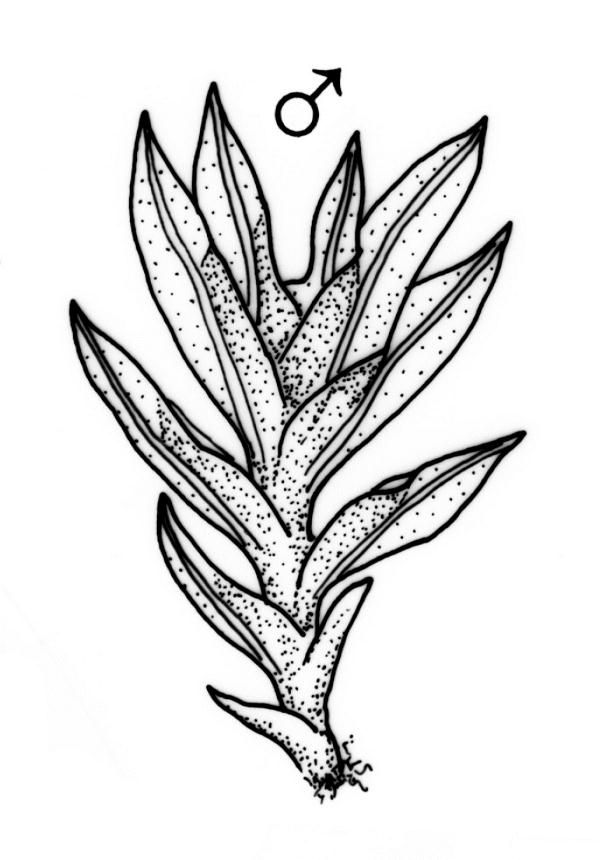
Plants heteromorphic, 1.5–5(–8) mm, pale to dark green, loosely or densely gregarious. Stems usually simple, rhizoids near base only. Leaves in 3–8(–15) pairs, patent, plane when moist, crispate when dry, oblong-lanceolate to linear-lanceolate, 0.5–1.0 × 0.10–0.25 mm; apex acute to acuminate, often cuspidate; laminae unistratose; vaginant laminae ⅔ leaf length, closed (or nearly so), but with minor lamina often deeply indented near its apex; dorsal lamina failing above, or reaching leaf insertion, tapered to the base; margins entire; marginal cells differentiated to form borders, well-defined on all laminae (var. curvatus), or poorly defined and incomplete or absent from dorsal and apical laminae (var. inclinabilis); cells of apical and dorsal laminae irregularly quadrate to hexagonal, smooth, non-bulging, thin-walled, (5–)8–12(–18) × (6–)7–11(–14) µm; cells of vaginant laminae longer, to 24 µm at mid lamina. Costa subpercurrent to long-excurrent, bryoides-type in cross-section.
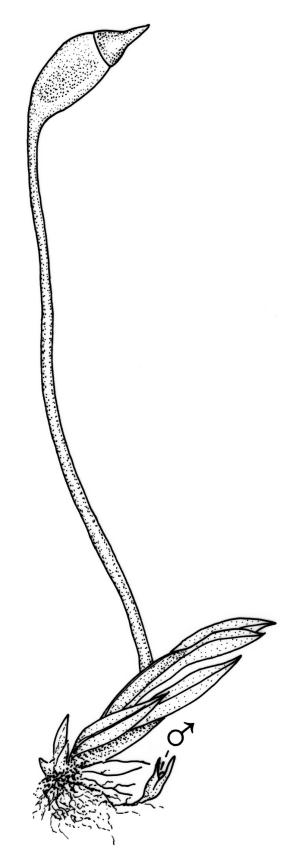
Autoicous or apparently dioicous. Perichaetia terminal; perichaetial leaves longer than vegetative, to 2.0 mm. Perigonia terminal on apparently independent, minute (to 2 mm) male plants, or bud-like and rhizautoicous. Setae yellow to orange-brown, 2.5–15 mm; capsules 0.5–1.0 mm, erect and symmetric to horizontal and arcuate; operculum rostrate from a conic base, with the beak erect to slightly inclined, ⅔ the length of theca. Peristome bryoides-type; teeth with long-columnar papillae on the adaxial trabeculae, 40–53 µm wide at base. Calyptra smooth, cucullate. Spores 12–16(–21) µm.
| Category | Number |
|---|---|
| Indigenous (Non-endemic) | 1 |
| Origin uncertain: Present in wild | 1 |
| Total | 2 |
The variability of capsule form and orientation in N.Z. material is considerable, ranging from symmetric and erect to highly asymmetric and horizontal; a similar variation has been noted throughout the species’ range (Pursell 1994). The distinction between var. curvatus and var. inclinabilis is made on the extent of the leaf borders, and this character is not correlated with capsule form.
The South African lectotype has shorter setae, but is otherwise similar to the N.Z. form that has ± erect capsules.
In F. curvatus, leaves that surround over-mature unfertilised archegonia have the form of vegetative leaves, which suggests that development of typical perichaetial leaf morphology requires fertilisation. Innermost perichaetial leaves at the base of a capsule are sometimes much smaller even than vegetative leaves, and have no meristematic cells remaining. Thus, after a period, the growth-promoting effect of fertilisation on the development of the surrounding leaves apparently ceases, and development of the innermost leaves is arrested.
It has been tentatively suggested (Beever 1999) that F. curvatus is adventive in N.Z., and may have moved from anthropogenic to natural habitats, where it is much less common. (e.g., on the soil among roots under a wind-thrown Rhopalostylis sapida in coastal forest on Taranga/Hen I. (J.E. Beever 65-37, AK 348402)). There is good evidence (Pursell 2006) that in California the species is adventive (see also below), and has spread from a c. 1948 arrival in the San Francisco Bay area. The high variability of this moss in N.Z. is an argument against its being adventive here, but the N.Z. population may have both indigenous and adventive components.