Class
Family
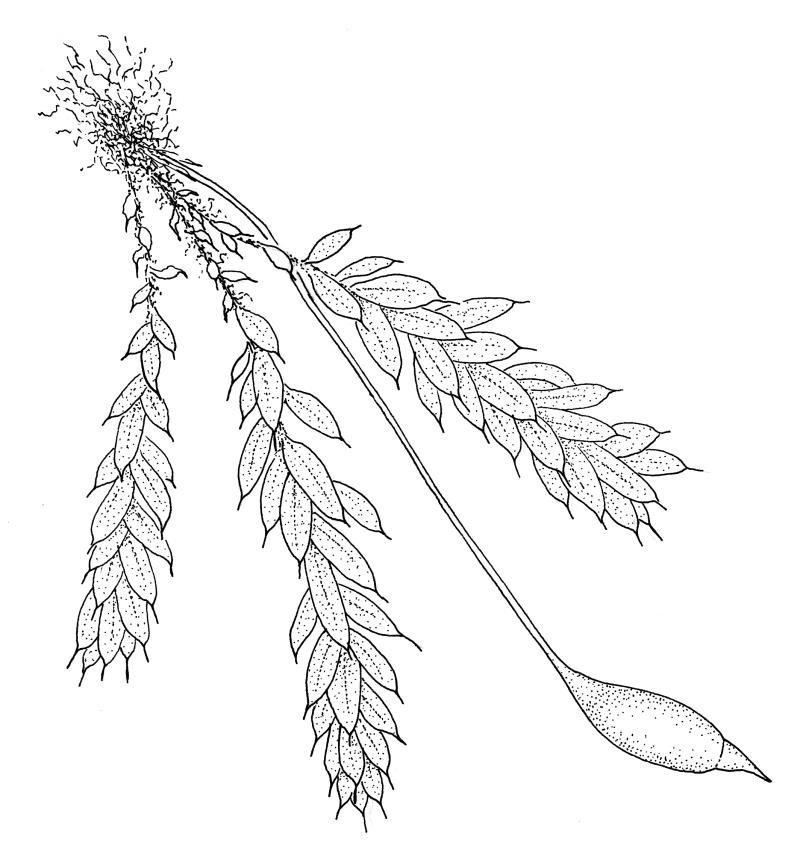
Plants small, glaucous, forming tufts, growing mostly on tree-fern caudices. Stems erect, branched at base, pale green or sometimes red-brown near base, beset at base with red-brown and papillose rhizoids. Leaves complanate or evenly distributed on shoots, erect-spreading, with a long, slender, and conspicuous hair-point, broadly elliptic to narrowly lanceolate, unbordered, crenulate or occasionally weakly dentate, reduced towards the stem base, with the lamina varying in width (in number of cells) between species. Upper laminal cells ± isodiametric or oblate, firm-walled, weakly to strongly mammillose, with an obscurely wrinkled cuticle (in N.Z. species), undifferentiated or becoming slightly larger towards the insertion; alar cells and marginal cells not differentiated; hair-point strongly developed, pale and conspicuous, persistent or sometimes caducous, sometimes consisting of a single cell for half or more of its length. Costa single, mostly percurrent and appearing to fuse with the hair-point or ending below the hair-point base (in the N.Z. species). Axillary gemmae absent (in the N.Z. species) or rarely present, many-celled and filamentous.
Rhizautoicous. Perichaetia at base of stem, red-brown, the leaves longer than vegetative leaves (in the N.Z. species). Perigonia bud-like, nidulant among basal rhizoids or rarely among leaf axils. Setae straight, smooth, pale brown; capsules erect or slightly inclined, ellipsoid, cylindric, or ovoid, pale red- or yellow-brown; exothecial cells firm-walled, smooth, isodiametric or rectangular; stomata few and difficult to observe, superficial (fide Tessler 2012); annulus weakly differentiated or apparently absent; operculum obliquely rostrate. Peristome single or absent in some species; exostome teeth mostly pale, strongly striate (in N.Z. species) or not, arising from a very low basal membrane (in the N.Z. species); endostome lacking. Calyptra cucullate. Spores spherical, smooth or slightly roughened.
Tessler (2012), in his recent revision of Hymenodon, recognised eight species in the genus, occurring in south-east Asia, Malesia, New Caledonia, Australia, N.Z., and tropical America. Koponen et al. (1986) have treated P.N.G. and western Melanesian species and included a key to the six species they recognised world-wide.
Six species are recorded by Tessler (2012) from the general area of Borneo south-east to New Caledonia. Among these six, H. angustifolius Sande Lac. seems a very distinctive species, but three others require further critical comparison. Hymenodon sericeum (Dozy & Molk.) Müll.Hal., H. tenellus Broth. & Paris, and H. chenianus Pócs may prove to be conspecific after further investigations. Combinations for two of these names have been proposed at the subspecies rank within H. pilifer, but these combinations seem inappropriate. The detailed consideration of the status of these three unconvincing species is beyond the scope of this treatment.
Shaw & Anderson (1986) investigated the nature of the peristome in Hymenodon sericeus and H. angustifolius, concluding that the single peristome in these two species is exostomal in nature. They confirmed the separation of the monotypic New Guinean Hymenodontopsis Herzog as distinct on the basis of it having an endostome. Earlier workers, including Brotherus (1924) and Koponen et al. (1986), considered the peristome of some or all species of Hymenodon to be endostomal in nature.
N.Z. material of H. pilifer is rhizautoicous. This corroborates tentative statements by Koponen et al. (1986) concerning the sexuality of H. sericeus and H. angustifolius (of P.N.G.) and observations by Tessler (2012) concerning the genus as a whole.
Recently Goffinet et al. (2009) placed Hymenodon (together with Leptotheca) in the Orthodontiaceae, following the molecular study of Bell et al. (2007). It is retained in this family here, albeit with reservations discussed above.
| Category | Number |
|---|---|
| Indigenous (Non-endemic) | 1 |
| Total | 1 |