Class
Subclass
Order
Family
Genus
Terrestrial or rupestral ferns. Rhizomes short and erect or rarely short-creeping, bearing scales. Rhizome scales narrowly ovate or ovate, 2–13 mm long, 0.5–2 mm wide, brown, clathrate. Fronds 25–425 mm long. Stipes 10–200 mm long, pale brown abaxially and proximally, green adaxially and distally, bearing abundant narrowly ovate or ovate scales with filiform apices. Rachises green, abundantly scaly, not winged or with a very narrow wing. Laminae usually 2-pinnate to 4-pinnate-pinnatifid, rarely 1-pinnate in smallest fronds, narrowly elliptic to broadly elliptic or ovate to broadly ovate, tapering gradually to the apex, 15–300 mm long, 8–200 mm wide, dark green on both surfaces, herbaceous, bearing scales, lacking hairs. Primary pinnae in 3–18 pairs below pinnatifid apex, not or only slightly overlapping; proximal pinnae narrowly ovate or ovate, or rarely broadly ovate in 1-pinnate fronds, those at mid-lamina ovate or narrowly ovate; the longest at or below the middle, 4–155 mm long, 2.5–50 mm wide, apices rounded or obtuse or acute or acuminate, bases stalked; costae of primary pinnae winged only in distal half. Secondary pinnae gradually decreasing in length along each primary pinna to the distal end; the longest proximal secondary pinnae ovate or elliptic or broadly ovate or suborbicular, 2.5–30 mm long, 2–10 mm wide, apices acute or obtuse or rounded, often partly or completely divided into tertiary segments, bases stalked; the distal secondary pinnae elliptic or narrowly oblong or linear, apices acute or obtuse, bases adnate. Tertiary pinnae elliptic or narrowly oblong or linear, 2–8 mm long, 0.3–3 mm wide, margins entire or divided, rarely divided into quaternary pinnae. Quaternary pinnae linear, up to 4 mm long and 0.3–0.5 mm wide, apices acute, margins entire, bases adnate. Sori submarginal on linear segments, away from margins on broader segments; indusia 1–4 mm long, straight; free margins of indusia entire. Mean spore size 31–37 μm long, 23–27 μm wide; perispores prominently winged and ridged.
Asplenium hookerianum is an extremely variable species. Two or more seemingly discrete morphologies can often be found growing together – one with rather broad, often rounded segments, another with very fine, linear segments. Both forms also vary in their degree of dissection from 2-pinnate in small plants to 4-pinnate in large plants.
Asplenium hookerianum is similar to small plants of A. gracillimum, but always lacks bulbils and generally has smaller laminae (15–300 mm long, 8–200 mm wide, cf. 60–800 mm long, 24–350 mm wide). Large fronds of A. hookerianum can be distinguished from small fronds of both A. bulbiferum and A. gracillimum by their clearly stalked secondary pinnae. Asplenium hookerianum is more difficult to distinguish from A. richardii, reflecting the fact that the latter is believed to be an autopolyploid derivative of the former (see Notes below). Asplenium richardii is a more robust plant and has pinnae that tend to overlap and spread in three dimensions, whereas A. hookerianum has two-dimensional fronds with pinnae that are well separated, at least in small fronds. The two species have different chromosome numbers and can be distinguished by spore size (44–49 × 32–36 μm in A. richardii cf. 31–37 × 23–27 μm in A. hookerianum). They are also partially ecologically separated, with A. richardii occurring at generally higher altitudes than A. hookerianum.
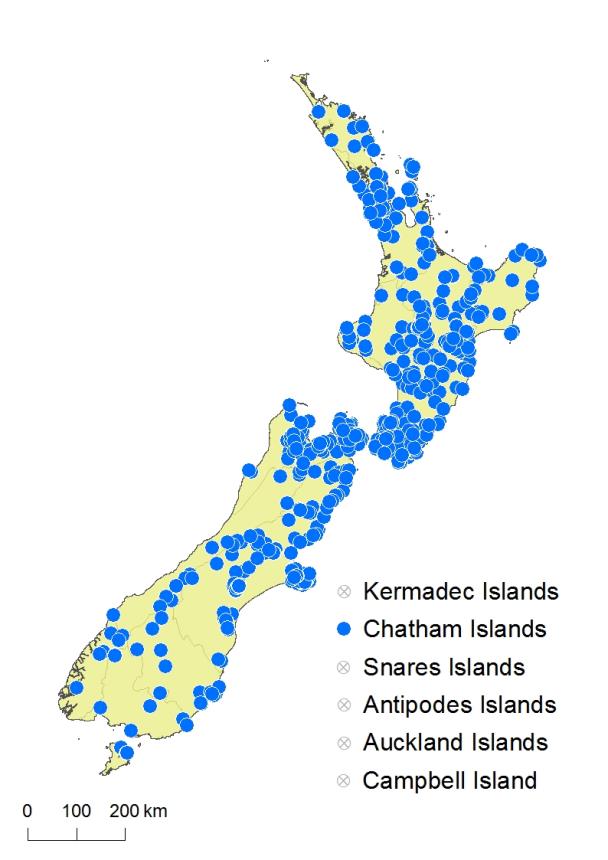
North Island: Northland, Auckland, Volcanic Plateau, Gisborne, Taranaki, Southern North Island.
South Island: Western Nelson, Sounds-Nelson, Marlborough, Westland, Canterbury, Otago, Southland, Fiordland.
Chatham Islands, Stewart Island.
Altitudinal range: 5–1375 m.
Asplenium hookerianum is widely distributed throughout lowland and montane forested areas of the North Island, from Kaitāia south. However, it is uncommon in Northland, inland Taranaki and the western Waikato. It grows from near sea level to over 1000 m on Mt Tongariro. In the South Island it is largely confined to lowland, montane and subalpine areas east of the main divide, growing from near sea level to 1375 m in north-west Nelson. However, it is scarce above 1000 m in both islands. It extends to Stewart Island and the Chatham Islands but is uncommon in both places.
Also Australia (Victoria, Tasmania).
Asplenium hookerianum is a terrestrial fern that occurs under open kauri, podocarp, broadleaved, and beech forest, in open kānuka, mānuka, and scrub, under Pinus and Cupressus macrocarpa, and among grass or in open pasture. It grows on well-drained soil, in shingle, on old sand-dunes, among rocks, in rock crevices and on rock faces, in sinkholes and under overhangs, on river terraces, banks and tracksides, among roots and tree bases, on upturned root plates and on dead trunks. It is found on a wide variety of rock types, including greywacke, scoria, schist, serpentine, limestone and marble.
There is evidence that A. hookerianum hybridises with A. appendiculatum subsp. appendiculatum, A. bulbiferum, A. flaccidum subsp. flaccidum (Brownsey 1977a), A. oblongifolium (Perrie & Brownsey 2004) and A. gracillimum (newly recorded here).
n = 72 (Brownlie 1958; Brownsey 1977b).
This taxon was first described by Raoul (1844) as Asplenium adiantoides. However, that name was illegitimate because of two earlier homonyms. It was subsequently described by Colenso (1845), who recognised broad-pinnuled and narrow-pinnuled forms as A. hookerianum and A. colensoi, respectively. Mettenius (1859) proposed A. raoulii as a replacement name for Raoul’s A. adiantoides, but in so doing he cited A. hookerianum and A. colensoi in the synonymy of A. raoulii var. minus and A. raoulii var. colensoi, and thus also in the synonymy of A. raoulii itself. The name A. raoulii is therefore nomenclaturally superfluous because either A. hookerianum or A. colensoi should have been used instead.
Because A. adiantoides Raoul and A. raoulii Mett. are illegitimate, the names A. adiantoides var. colensoi (Colenso) Hook.f., A. adiantoides var. hookerianum (Colenso) Hook.f., A. adiantoides var. minus Hook.f., A. raoulii var. colensoi (Colenso) Mett., and A. raoulii var. minus (Hook.f.) Mett. are all incorrectly formed and cannot be used, even though the varietal epithets themselves are legitimate.
A minor mystery surrounds Colenso’s apparent defiance of protocol in naming A. colensoi after himself. Earp (2018) has summarised this, but concludes that any explanation is necessarily speculative.
Asplenium rotundifolium was described by Ettingshausen (1864) from material collected "in Nova Zeelandia", but without any other details, such as collector or date. As far as is known, Ettingshausen never visited New Zealand, so the name is unlikely to have been based on his own collections. In the protologue, "Tarachia rotundifolia Presl" is cited in synonymy, but that name has never been published. A specimen in W is labelled "Tarachia rotundifolia Presl", was collected by Hügel in New Zealand, and, in the absence of any other original material is the presumed holotype of Asplenium rotundifolium. The material is clearly A. hookerianum.
The broad-pinnuled and narrow-pinnuled forms of A. hookerianum have been accepted as varieties in all New Zealand Floras since 1867 (Hooker 1864; Cheeseman 1906, 1925; Allan 1961), or as full species by other authors (e.g. Crookes 1963). However, following Brownsey (1977b), recent treatments have accepted only A. hookerianum as one polymorphic species (Brownsey et al. 1985; Brownsey & Smith-Dodsworth 2000). Using AFLP DNA-fingerprinting and chloroplast DNA sequencing Perrie & Brownsey (2005) showed that genetic variation in A. hookerianum is not concordant with pinnule morphology, and that plants from the same site, irrespective of their pinnule morphology, are generally more closely related to each other than they are to plants with the same pinnule morphology from other sites. They concluded that there was no justification for recognising the narrow-pinnuled form as var. colensoi.
Shepherd et al. (2007, 2009) showed that A. hookerianum has extensive chloroplast DNA variation, with at least 25 different haplotypes, suggesting that it is an old lineage which survived the last glacial period in widespread and possibly large populations. This, together with its preference for dry habitats, may account for the fact that A. hookerianum is implicated in more polyploid lineages than any other New Zealand species (Shepherd et al. 2008); it may also account for at least some of the morphological variation. Analysis of low-copy nuclear DNA sequences suggests that A. hookerianum is one tetraploid parent of four allopolyploid species, A. cimmeriorum, A. gracillimum, A. lyallii, and A. appendiculatum, and the sole parent of the autopolyploid species A. richardii.
In Australia A. hookerianum is known from two small populations in Victoria and two in Tasmania (Brownsey 1998; Perrie et al. 2010). In contrast to New Zealand populations, only two haplotypes have been recorded (Perrie et al. 2010). The pattern of haplotype variation in Australia is consistent with two independent dispersals of A. hookerianum from New Zealand to Australia, a finding that is unique in trans-Tasman dispersal of vascular plants.
Shepherd et al. (2009) demonstrated the presence of two haplotypes in populations of A. hookerianum on the Chatham Islands. Each was more closely related to haplotypes found in mainland New Zealand than they were to each other. Shepherd et al. (2009) concluded that populations of A. hookerianum on the Chathams are derived from at least two long-distance dispersal events, indicating that the species has dispersed over a significant distance both east and west.