Class
Subclass
Order
Family
Genus
Rhizome scales acicular or narrowly ovate with filiform apices, 5–30 mm long, 0.5–2 mm wide, clathrate. Fronds 75–1560 mm long. Stipes 15–440 mm long, bearing scattered narrowly ovate scales with filiform apices. Laminae usually 1-pinnate-pinnatifid, rarely 2-pinnate-pinnatifid, ovate or elliptic, 60–1280 mm long, 20–250 mm wide, limp and pendulous. Primary pinnae in 2–26 pairs below pinnatifid apex, not overlapping; the proximal pinnae and those at mid-lamina linear to narrowly ovate, or rarely ovate in primary pinnae with an enlarged basal acroscopic secondary pinna; the longest 20–340 mm long, 3–24 mm wide (excluding basal acroscopic pinnule), apices drawn out into undivided narrowly acuminate segments 6–50 mm long. Secondary segments usually decreasing slightly in length both distally and proximally along each primary pinna, but rarely the basal acroscopic one extended and further divided; the longest secondary segments 2–20 mm long, 0.75–2 mm wide; rarely the basal acroscopic secondary pinna extended up to 100 mm long and 8 mm wide, undivided or bifid or divided into linear or oblong tertiary segments. Indusia 2–13 mm long. Mean spore size 35–46 μm long, 21–30 μm wide; perispores with a prominent wing but lacking ridges.
Note: excludes collections from the Kermadec Islands. The values given above are from measurable herbarium specimens. Larger fronds are known to occur in the wild. CHR 23766 comprises parts of a frond that was said to measure 7 ft 4 in (c. 2235 mm) long.
Asplenium flaccidum subsp. flaccidum is typically an epiphytic forest plant growing on tree trunks or hanging from Astelia clumps. It has limp, pendulous, leathery fronds which are usually 1‑pinnate-pinnatifid, up to 1.5 m in length, and with the primary pinnae divided almost to the midrib into narrow segments, which bear submarginal, elongated sori. The secondary segments on each primary pinna are usually more or less equal in length, decreasing slightly in size both distally and proximally. However, some populations, especially in the western part of the South Island, have laminae in which the basal acroscopic segment is much longer than the adjacent ones. Sometimes plants of A. flaccidum subsp. flaccidum grow on the ground, especially in rocky scrub and under introduced pine plantations. Such plants have much shorter fronds, but they are still rather limp and pendulous.
Asplenium flaccidum subsp. flaccidum is sometimes confused with A. appendiculatum but can be distinguished by its usually epiphytic and pendulous habit, whereas A. appendiculatum grows terrestrially, producing upright fronds that are also more divided than in A. flaccidum. The latter is also tetraploid rather than octoploid, with smaller and less prominently ridged spores.
Subsp. flaccidum can usually be distinguished from subsp. haurakiense by its habitat and growth form. Subsp. haurakiense almost always grows on the ground or on rocks, occasionally as a low epiphyte on Metrosideros excelsa, in close proximity to the sea. It is confined to the east coast of Northland and islands of the Hauraki Gulf, extending south to the Bay of Plenty. Fronds grow upright, are firmer than in subsp. flaccidum, frequently (but not invariably) have an enlarged basal acroscopic segment on each primary pinna (similar to that sometimes seen in subsp. flaccidum), and have larger spores (40–49 × 26–33 μm, cf. 35–46 × 21–30 μm, excluding populations from the Kermadec Islands). They also have a slightly different perispore pattern, with less pronounced wings (see Brownsey 1977b; Large & Braggins 1991). The stipe scales are ovate, lack filiform apices, and have thicker cell walls in contrast to the narrower, more drawn out stipe scales of subsp. flaccidum.
On the east coast of Northland and on the Hauraki Gulf islands there are relatively few places where the two taxa grow abundantly in close proximity to each other. However, in areas where the two subspecies do occur together (e.g. Poor Knights Islands, Little Barrier Island), it is sometimes difficult to distinguish individual plants, and there is significant overlap in spore size. Nevertheless, most plants can be assigned to one or other of the taxa; plants with spores less than 40 μm long are likely to be subsp. flaccidum, and those with spores wider than 26 μm are usually subsp. haurakiense.
The situation on the Kermadec Islands, particularly on Raoul Island, is rather different. On this larger island there is more extensive forest and a long rocky coastline providing abundant habitat for both taxa. A few collections at either end of the morphological spectrum can be assigned to one or other taxon, but the majority of plants appear to be intermediate in morphology, or have a form not seen elsewhere in New Zealand, and are not strongly correlated with habitat. Thus epiphytic forms often have extended basal acroscopic segments, and terrestrial forms sometimes almost completely lack them. Spore size in Kermadec material also varies hugely, with an even greater range than recorded for the two taxa elsewhere in New Zealand (30–49 × 21–30 μm long, cf. 35–49 × 21–33 μm), but size is not correlated with habitat as it is the northern North Island. As noted by Sykes & West (1996), it appears that on the Kermadec Islands the two forms are less clearly distinguished and most plants can only be identified as Asplenium flaccidum sensu lato.
No plants with aborted spores have been detected amongst over 75 individual collections examined from throughout its range, including many that are of intermediate morphology. This, together with the wide range of morphology observed on the Kermadec Islands, uncorrelated with habitat, suggests that the two forms are best treated as susbspecies rather than species. It would be possible to test this hypothesis by an in-depth analysis of genetic variation in areas where the two forms regularly occur together (e.g. Kermadec Islands, Poor Knights Islands, Little Barrier Island).
North Island: Northland, Auckland, Volcanic Plateau, Gisborne, Taranaki, Southern North Island.
South Island: Western Nelson, Sounds-Nelson, Marlborough, Westland, Canterbury, Otago, Southland, Fiordland.
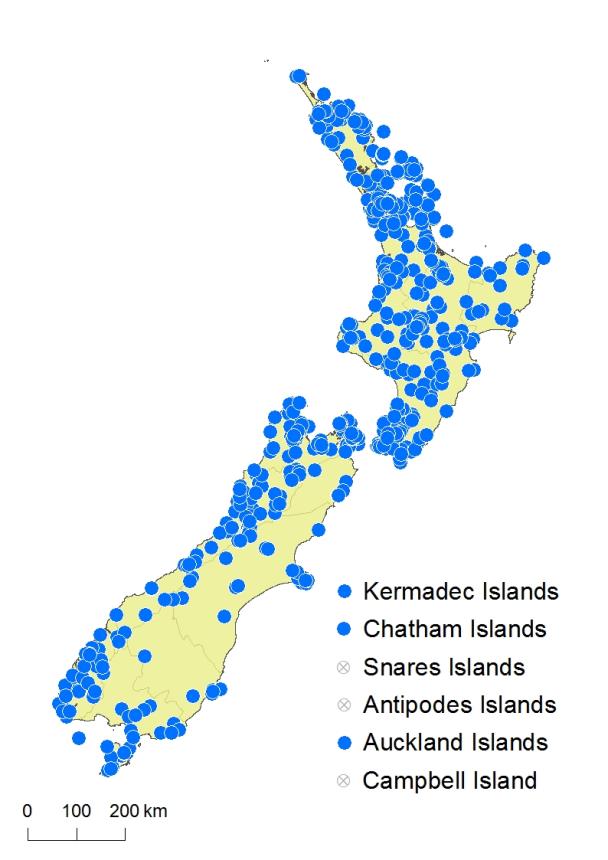
Kermadec Islands, Chatham Islands, Solander Island, Stewart Island, Auckland Islands.
Altitudinal range: 0–1525 m.
Asplenium flaccidum subsp. flaccidum occurs on the Kermadec Islands (e.g. AK 352152) and is widely distributed throughout lowland and montane forested areas of the North Island. It grows from near sea level to 1100 m in the Kaimanawa Ranges. In the South Island it occurs mostly frequently in lowland to subalpine areas west of the main divide, growing from near sea level to over 1500 m on Mt Arthur, with only scattered populations on the east coast and in inland Canterbury. It extends to the Chatham Islands, Solander Island, Stewart Island and Auckland Islands. It was erroneously recorded from the Snares Islands by Brownsey & Smith-Dodsworth (2000).
Also Australia (Queensland, New South Wales, Victoria, Tasmania).
Asplenium flaccidum subsp. flaccidum usually grows as an epiphyte in kauri, podocarp, beech and broadleaved forest, and in mānuka, kānuka and low scrub. It has been recorded growing on Cyathea cunninghamii, C. dealbata, C. medullaris, C. smithii, Dicksonia squarrosa, Beilschmiedia tawa, Carex secta, Carpodetus serratus, Cordyline australis, Coriaria arborea, Corynocarpus laevigatus, Dacrydium cupressinum, Fuchsia excorticata, Griselinia littoralis, Hedycarya arborea, Knightia excelsa, Kunzea ericoides, Leptospermum scoparium, Melicytus ramiflorus, Metrosideros spp., Nestegis montana, Olearia colensoi, Pinus sp., Podocarpus totara, Prumnopitys ferruginea, Rhopalostylis sapida, Salix sp., Streblus banksii, Vitex lucens, Weinmannia racemosa, species of Nothofagaceae, and from clumps of Astelia, as well as on aerial roots, rotten trunks and stumps. It sometimes also grows terrestrially on cliffs, rocks and scoria, in crevices, on mossy banks, stream banks, limestone and marble, thermally heated soil, under Cupressus macrocarpa and Pinus, and rarely on sand dunes under marram grass.
There is evidence that A. flaccidum subsp. flaccidum hybridises with A. appendiculatum subsp. appendiculatum, A. appendiculatum subsp. maritimum, A. bulbiferum, A. hookerianum, A. oblongifolium, A. obtusatum, A. scleroprium (Brownsey 1977a), A. gracillimum, A. lyallii (de Lange et al. 2011), A. lepidotum (Perrie & Brownsey 2016) and A. chathamense (newly recorded here). The latter combination is based on specimens (AK 300982 and WELT P022277) originally cited by de Lange et al. (2011) as A. flaccidum × lyallii.
n = 72 (Brownlie 1954; Brownsey 1977b).