Class
Family
Genus
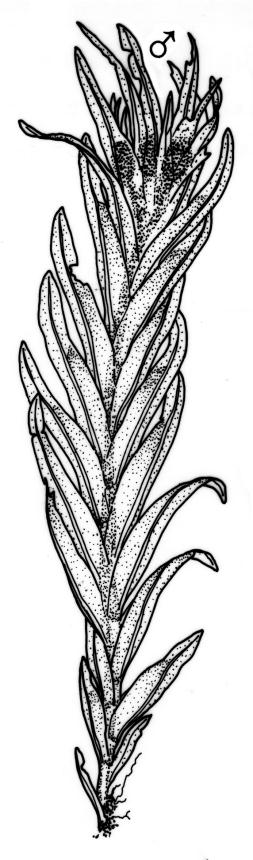
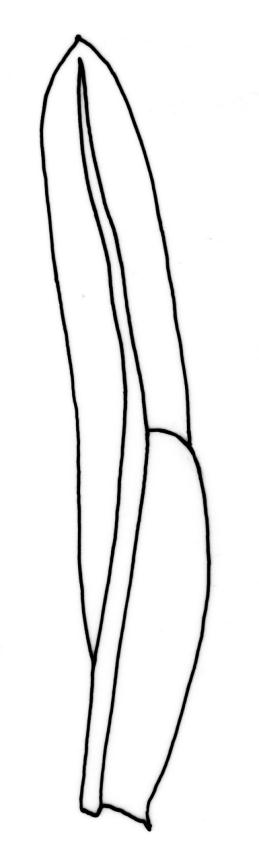
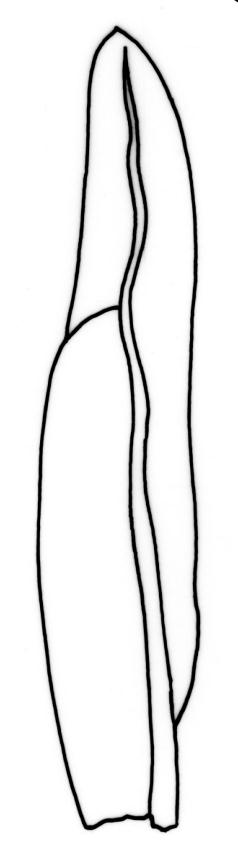
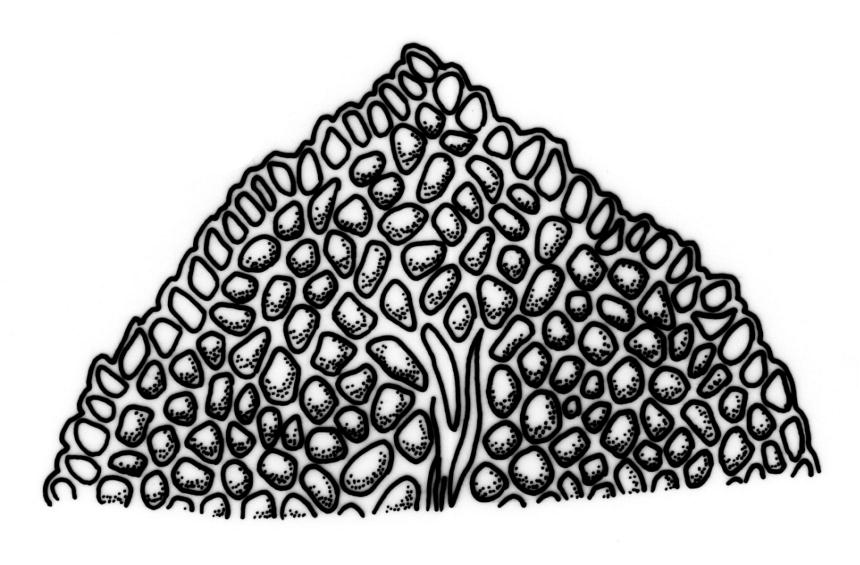
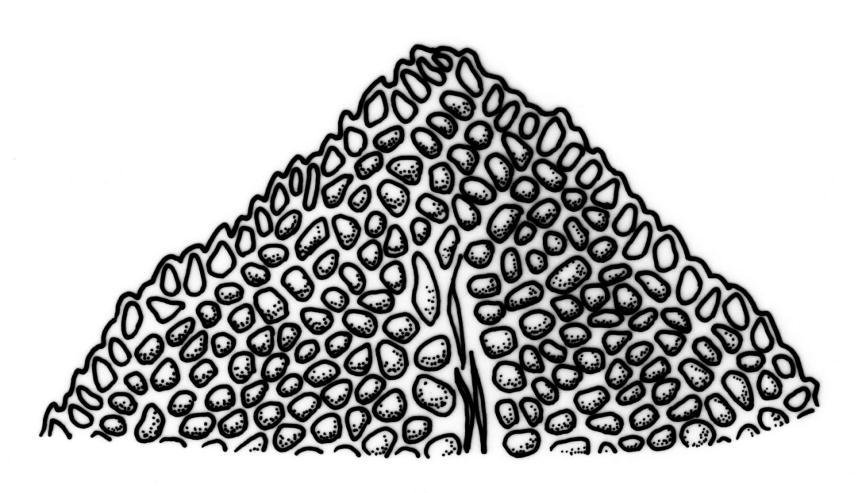
Plants 5–20(–65 in aquatic forms) mm, yellow-green to dark green, bronze or black below, densely gregarious. Stems occasionally branched, with rhizoids at branch bases. Leaves in 10–25 pairs, scarcely overlapping at mid stem, patent, decurved when moist, apices often strongly inrolled towards the substrate when dry, oblong-lingulate, 2.0–4.0 × 0.3–0.7 mm; apex obtuse; laminae unistratose; vaginant laminae ½–¾ leaf length, open or nearly open, with the minor lamina often rounded at apex; dorsal lamina often failing above leaf insertion, tapered to its base; margins serrulate on dorsal and apical laminae, irregularly so near the leaf apex, serrulate in distal region of vaginant laminae, ± entire in supra-basal region of vaginant laminae, with marginal cells there often elongated obliquely or parallel to the margin; cells of apical and dorsal laminae irregularly rounded-hexagonal, smooth, bulging, with medium-thick walls (6–)7–11(–15) × (6–)7–11(–15) µm. Costa failing 4–8 cells below the leaf apex, modified oblongifolius-type in cross-section (epidermal cells with larger lumina than those of the stereid cells beneath).
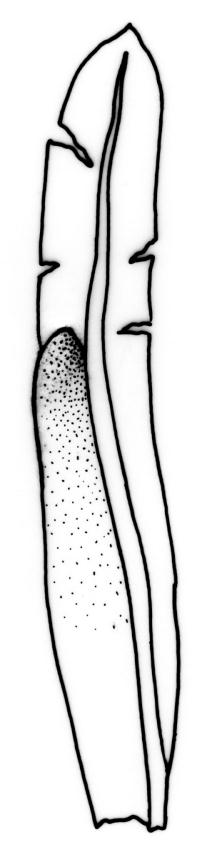
Dioicous. Perichaetia terminal on tall plants; perichaetial leaves longer than vegetative, with acute apices. Perigonia terminal on tall plants. Setae yellow to orange-brown, stout, arcuate, scarcely twisted when dry, 3–5 mm; capsules inclined, ± symmetric, 0.75–1.50 mm; operculum obliquely rostrate or erect-rostrate from a conic base, ½ to equal the length of theca. Peristome fasciculatus-type; teeth 65–100(–110) µm wide at base. Calyptra smooth, cucullate. Spores 14–19 µm.
Wilson 1854, pl. 84, fig. 1 (as F. ligulatus); Scott & Stone 1976, pls 7– 9; Catcheside 1980, fig. 22; Magill 1981, figs 15, 13–18; Crum & Anderson 1981, fig. 32, D–H; Beever & Stone 1998, figs 2, b, 3–4, 5, g, 7, b; Beever et al. 2002, p. 16, figs 1–6; Bruggeman-Nannenga & Arts 2010, fig. 8; Stone & Catcheside 2012.
The appropriate placement of F. asplenioides in a subgeneric classification is problematic. The species has been placed in sect. Amblyothallia Müll.Hal., both traditionally by Brotherus (1924) and more recently in numerous publications (Bruggeman-Nannenga & Berendsen 1990; Beever 1999; Pursell & Bruggeman-Nannenga 2004; Pursell 2007). Alternatively, the species has been placed in sect. Serridium Müll.Hal., a move supported by Allen (1980), following his extensive SEM studies of Fissidens peristomes. The peristome of F. asplenioides is, however, unusual in having flat, scarcely twisted filaments, which are coarsely papillose in their most distal parts. Bruggeman-Nannenga & Berendsen (1990), in another major study of Fissidens peristomes, named this the fasciculatus-type. They noted that it was the only peristome type that was not restricted to the Fissidentaceae: it is found also in the Dicranaceae.
In the opinion of the present author, F. asplenioides is misplaced with its traditional allies, which in the N.Z. context comprise F. capitatus, F. hyophilus, F. oblongifolius, and F. pallidus. It differs from those species not only in its peristome, but also in its costal structure: seen in cross-section, the epidermal cells have significantly larger lumina than do the stereid cells they overlie, and the tissues of the vaginant laminae, where they abut on the costa, are distinctive in being complex.
Suzuki & Iwatsuki (2007) noted that F. asplenioides had been selected as the lectotype species of sect. Amblyothallia by Bruggeman-Nannenga et al. (1994). They therefore established a new subgenus (Neoamblyothallia Tad.Suzuki & Z.Iwats.) to accommodate the other members of former sect. Amblyothallia, an arrangement that has considerable merit.
Fissidens asplenioides is easily confused with F. oblongifolius. Sterile material may be unequivocally distinguished under the microscope by examination of the supra-basal region of the vaginant laminae. In F. asplenioides this margin is entire, with marginal cells longer than wide, whereas in F. oblongifolius (and the related F. capitatus and F. hyophilus) this margin is crenulate from the bulging periclinal walls of oblate marginal cells. When plants are fruiting, the short and stout setae, nearly symmetric capsules, and fasciculatus-type peristome of F. asplenioides are diagnostic. The peristome can be recognised with a hand-lens, as the filaments are smoothly incurved when dry, rather than irregularly splayed, as in all other N.Z. Fissidens species (F. taylorii var. sainsburyanus, a much smaller plant, excepted). A further diagnostic feature of F. asplenioides is an open and rounded apex on the minor vaginant lamina of some leaves.
K; NI: N Auckland, including offshore islands (TK, PK, LB, GB, RT), S Auckland, Gisborne, Hawke’s Bay, Taranaki, Wellington (including KA); SI: Nelson (including D’U), Marlborough, Canterbury, Westland, Otago, Southland; St; Ch; A.
Anomalous. Tasmania*, mainland Australia*, southern Africa*, Chile*. Reported from south-eastern North America including Mexico, and south-east Asia by Crum & Anderson (1981) and considered to be "essentially pantropical" by Pursell (1994).
A very common species, on soil or rock, predominantly vertical or sloping, in a wide range of vegetation types, including indigenous and exotic forests, scrub, fernland, rough pasture and shaded lawns in urban parks. This species has probably become much more widespread with the advent of human disturbance in N.Z. It tolerates a wide range of insolation, and is characteristically more yellow in exposed sites. It occurs in both dry and mesic sites, may be intermittently submerged, or it may even be truly aquatic. Fissidens asplenioides is the most common species of the genus in N.Z. on vegetated roadside banks, lake margins, and on stream banks and emergent boulders in streams. When subject to periodic flooding, F. asplenioides often forms extensive, pure, and silt-impregnated swards. It is commonly found on wet rock in the splash zone of waterfalls and occasionally is the dominant species in coastal seeps. It is rarely found submerged in streams, but has been collected from below 10 m at the bottom of several South I. lakes (de Winton & Beever 2004). Fissidens asplenioides is often found associated with other species of the genus: F. blechnoides, F. curvatus, F. dealbatus, F. linearis, F. pallidus, F. taylorii, and F. tenellus have all been co-collected. Other associated mosses include Campylopus clavatus, C. introflexus, Catagonium nitens, Chrysoblastella chilensis, Didymodon australasiae, Distichophyllum pulchellum, Ditrichum difficile, Hypnum cupressiforme, Philonotis tenuis, Pyrrhobryum bifarium, Sematophyllum jolliffii, Thuidiopsis furfurosa, Tridontium tasmanicum, and Weissia controversa. This diverse array of associated species is indicative of the wide range of habitats occupied by F. asplenioides.
Records range from sea level to c. 900 m on Taranaki/Mt Egmont.
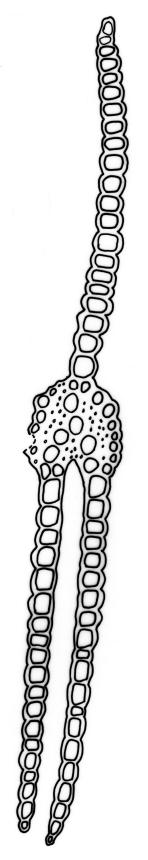
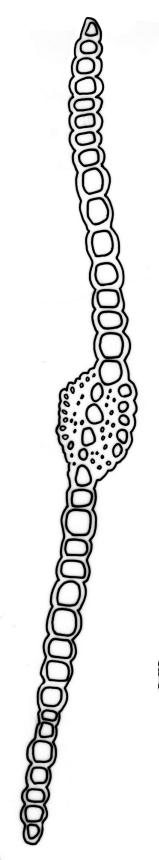
Pursell & Bruggeman-Nannenga (2004) reported that in F. asplenioides "the [axillary hyaline] nodules can at times be large"; such nodules have not been seen by the present author in N.Z. material. The only N.Z. taxon of Fissidens in which axillary nodules are conspicuous is F. crispulus var. robinsonii.
An account of F. asplenioides in N.Z. was given by Beever & Stone (1998).