Class
Subclass
Order
Family
Genus
Terrestrial or rupestral ferns. Rhizomes short-creeping, up to 70 mm long (in herbarium specimens), 3–5 mm diameter, bearing scales. Rhizome scales narrowly ovate or ovate, 4–10 mm long, 0.5–2 mm wide, blackish-brown, clathrate. Fronds 45–370 mm long. Stipes 20–150 mm long, yellow-brown or green, bearing ovate scales with short filiform projections throughout. Rachises green, scaly. Laminae 1-pinnate-pinnatifid to deeply 2-pinnate-pinnatifid, narrowly triangular or narrowly ovate or ovate, tapering gradually to the apex, 25–220 mm long, 10–78 mm wide, mid- to dark green on both surfaces, herbaceous, bearing scattered scales, lacking hairs. Primary pinnae in 3–18 pairs below pinnatifid apex, not or only slightly overlapping; proximal pinnae ovate, those at mid-lamina ovate or narrowly ovate; the longest below the middle, 7–55 mm long, 3–17 mm wide, apices obtuse, bases stalked. Secondary pinnae gradually decreasing in length along each primary pinna to the distal end, the basal acroscopic one the largest; the longest proximal secondary pinnae elliptic or ovate, 5–13 mm long, 1–7 mm wide, apices obtuse, incised or partly divided into tertiary segments, bases short-stalked or sessile; the distal secondary pinnae oblong, apices obtuse, bases adnate. Sori away from margin; indusia 1.5–3.5 mm long, straight; free margins of indusia entire or irregularly incised. Mean spore size 44–52 μm long, 33–39 μm wide; perispores with anastomosing wings and low tuberculae in the lacunae.
Asplenium cimmeriorum is morphologically similar to A. bulbiferum, A. gracillimum, and A. hookerianum, but differs from all three in having a short-creeping rhizome. It has generally smaller fronds than either A. bulbiferum or A. gracillimum, and lacks the bulbils of those species. It has a slightly narrower frond than A. hookerianum, and is octoploid rather than tetraploid, with larger spores (44–52 × 33–39 μm cf. 31–37 × 23–27 μm). Its habitat is also distinctive.
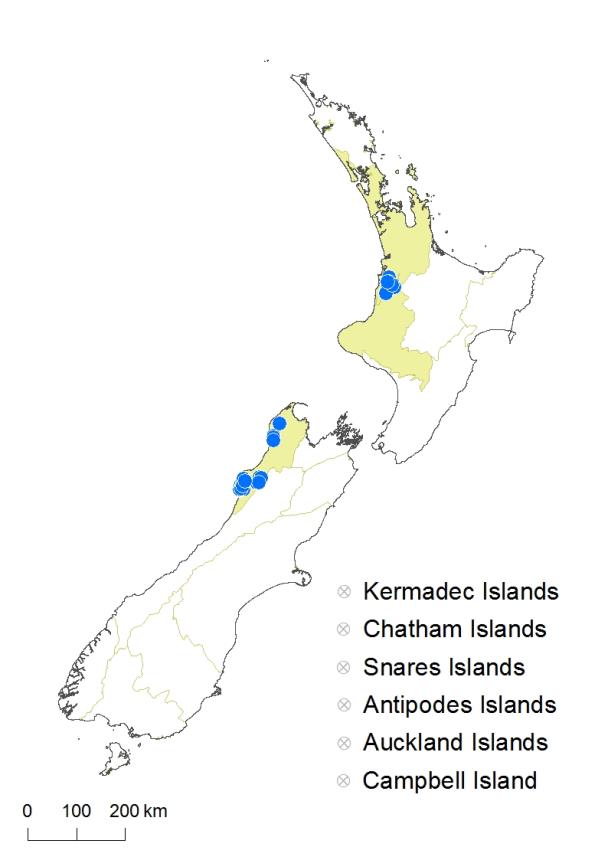
North Island: Auckland, Taranaki.
South Island: Western Nelson.
Altitudinal range: 5–620 m.
In the North Island Asplenium cimmeriorum occurs only in the Waitomo area of western Waikato, where it grows from 160 m to 460 m. In the South Island it occurs in north-west Nelson and north Westland from Gouland Downs to Punakāiki, growing from near sea level up to 620 m on the Heaphy Track. There are also occasional records from the Inangahua and Buller Valleys.
Asplenium cimmeriorum is largely confined to dark, calcareous sites in high-rainfall, lowland to montane areas (Brownsey & de Lange 1997). It grows most frequently at limestone cave entrances, under overhangs, on cave walls and ceilings or associated rockfalls, or sometimes in deeper caves near the limit of daylight penetration. In the South Island, plants also extend out into surrounding forest, growing on rotting logs, amongst tree roots, or on limestone boulders and rock faces.
Asplenium cimmeriorum was given a conservation status of Naturally Uncommon by de Lange et al. (2013).
There is evidence that A. cimmeriorum hybridises with A. bulbiferum (Brownsey & de Lange 1997).
n = c. 144 (Brownsey & de Lange 1997).
DNA analyses strongly suggest that A. cimmeriorum is an allopolyploid species derived from A. bulbiferum and A. hookerianum, and that the populations from the North and South Island have independent polyploid origins (Shepherd et al. 2008; Perrie et al. 2010).