Class
Classification
Subordinate Taxa
Nomenclature
Scientific Name:
Funariaceae Schwägr., Sp. Musc. Frond. 5(1), 43 (1830) – as Funariae
Type Taxon:
Description
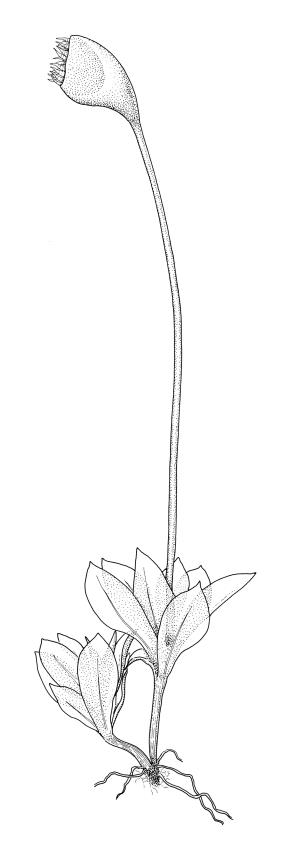
Plants minute to medium-sized, mostly pale green, lacking a persistent protonema, gregarious on soil. Stems branching by subperichaetial innovation, occasionally unbranched, or forked, not forming "rhizomes", usually red-brown, in cross-section with a central strand, 1–4 thick-walled cortical cell layers and a hyaloderm, beset with smooth, usually red-brown rhizoids; rarely with tubers. Leaves usually larger and more crowded above, lax, concave or less commonly plane, erect-spreading or rarely erect, usually obovate, less often elliptic, lanceolate, or subulate, entire or serrate by projecting cells, or very rarely ciliate, acuminate to obtuse; laminal cells large, thin-walled, and smooth, oblong-hexagonal or oblong, usually thinner-walled and more oblong in lower portion of leaf, occasionally elongate at margins to form a border; costa single, usually strong, protruding on abaxial surface, nearly always with a small (rarely large) central stereid group surrounded by one abaxial and one or two adaxial layers of larger cells. Axillary hairs always present, with long-cylindric terminal cells.
Mostly autoicous, less commonly paroicous, synoicous, or polygamous. Perigonia terminal, usually single, with multicellular paraphyses with globose or pyriform yellow terminal cells ("funariaceous"). Perichaetial shoot usually arising by subperigonial innovation and overtopping the perigonium; perichaetia terminal, lacking differentiated paraphyses. Capsules immersed to long-exserted, erect or curved, often strongly asymmetric, operculate or inoperculate, usually with a distinct neck; setae elongate or very short; stomata restricted to neck, consisting of an elongate pore in a single guard cell ("funariaceous"), immersed or superficial; exothecial cells thick- or thin-walled, the radial walls sometimes cuneate in cross-section; annulus present or absent. Peristome double, single, rudimentary, or absent; exostome teeth, if present, sigmoid or straight, sometimes apically fused as a latticed disc, papillose-striolate or striate, trabeculate on adaxial surface; endostome if present coherent at base, with or without a basal membrane, segments (if present) opposite the teeth, acute or irregular, papillose-striolate or papillose; cilia lacking. Calyptra deciduous or persistent, usually smooth, papillose; mitrate or cucullate, usually long-rostrate and inflated at base, rarely angled or pleated. Spores small to very large.
Taxonomy
The Funariaceae are a medium-sized family of cosmopolitan distribution. Six genera and 12 species are accepted from New Zealand. Fife (1985) recognised 13 genera worldwide, of which Entosthodon is the largest. Virtually without exception its members occupy bare soil, usually but not always with little humic material. In tropical regions they predominantly occur at higher elevations, although a few genera (mostly those of simple sporophyte morphology) occur in periodically flooded lowland tropical areas. The plants are short-lived, but often overwintering in temperate regions. New Zealand representatives often over-winter in the "spear" stage.
The Australian species were reviewed by Fife & Seppelt (2001).
The family is characterised by having large, thin-walled, and smooth, mostly oblong-hexagonal laminal cells, which are clearly visible under a hand-lens. The gametophytes in the family exhibit only modest variability, while the sporophytes are highly variable and provide most of the morphological features traditionally used to define genera and, in some instances, species. The family includes genera with long-exserted capsules with highly developed double peristomes, reduced peristomes, gymnostomous and operculate capsules, and inoperculate and sessile capsules.
Since von Wettstein’s (1932) pioneering studies on hybridisation in the family, the members of the Funariaceae, particularly Funaria hygrometrica and Physcomitrella patens, have been important experimental organisms. More recently the family has been the object of intense molecular and developmental study, and its members, particularly Physcomitrella patens, have been widely used in studies of gene expression; these studies have given rise to a large body of specialised literature (Rensing et al. 2008; Lang et al. 2008; McDaniel et al. 2010). Physcomitrella patens is the first bryophyte to have its genome completely sequenced and can legitimately be considered the Arabidopsis of the bryophytes.
The traditional interpretation of the family as a reduction series (in terms of sporophyte complexity) has been recently challenged by gene sequencing techniques and attempts at phylogenetic reconstruction. Homoplasy (convergent evolution) has been convincingly shown to play a major role in the evolution of the funariaceous sporophyte (Liu et al. 2012) and at least three genera (sensuFife 1985) (Physcomitrium, Entosthodon, and Physcomitrella) have been hypothesised as polyphyletic.
The resurrection of the genus Physcomitridium (Goffinet & Buck 2011; Hooper et al. 2010) was initially argued for using data derived from four nuclear gene loci (McDaniel et al. (2010)) and has subsequently received extremely strong corroboration by Liu et al. (2012), who used 10 gene loci from nuclear, chloroplast, and mitochondrial genomes to outline relationships in the Funariaceae. The genus Physcomitridium is accepted here. Accordingly, Physcomitrella is excluded from the N.Z. flora.
The generic circumscriptions of Entosthodon and Physcomitrium presented in this Flora are likely to be modified in future when consensus is reached concerning the "backbone" phylogenetic structure of the Funariaceae.
Material of Funariaceae is rarely collected without sporophytes, and this is reflected in the strong emphasis on sporophyte characters in the following key to genera. Indeed there are few, if any, reliable gametophytic features, even at a regional level, that permit the confident distinction of genera as they are circumscribed here. Because of the high level of autoicy, capsules are rarely completely absent from collections. Several species of N.Z. Funariaceae, however, do have distinctive gametophyte features (e.g., overall leaf shape, the nature of the leaf apex, the strength of the costa, the presence of differentiated margin cells and toothing, rhizoid colour), which would permit their recognition if sterile. Ironically, the two most widespread and frequent genera, Funaria and Physcomitrium, are perhaps the most difficult to confidently separate using gametophyte characters. The preference of Funaria hygrometrica for a burnt soil substrate can sometimes help to distinguish them if the plants are completely sterile, but it also often grows on unburnt soils.
Key
| 1 | Capsules clearly immersed, lacking a peristome; a functional operculum either present or absent; plants extremely rare and known only from North I. (N & S Auckland and Hawke’s Bay L.D.) | 2 |
| 1' | Capsules exserted (either weakly or strongly), with or without a peristome; a functional operculum present (except in Bryobeckettia and then capsules only weakly exserted); plants more common and often widespread | 4 |
| 2 | Capsules lacking a functional operculum; capsules globose, lacking a neck, often disintegrating due to extreme delicacy of the exothecium (and then leaving an exposed mass of spores); exothecial cells at mid urn extremely thin-walled, hyaline, not collenchymatous | Physcomitridium |
| 2' | Capsules with a functional operculum; capsules neither globose nor disintegrating due to the delicacy of the exothecium; exothecial cells firmer-walled, pigmented or hyaline, sometimes weakly collenchymatous | 3 |
| 3 | Calyptra mitrate, with 8 conspicuous radial pleats; mouth equal or slightly less than the diameter of capsule; spores ellipsoid, 40–110 μm in greater diam., yellow-brown, often appearing smooth under light microscope, becoming reticulate at maturity; plants known from a single Hawke’s Bay L.D. locality | Goniomitrium |
| 3' | Calyptra mitrate but lacking 8 radial pleats; mouth c. ⅓ the diameter of the capsule; spores spherical to subreniform, <50 µm in greater diam., red-brown, conspicuously spinose; plants known from N & S Auckland L.D. | Physcomitrium (pusillum) |
| 4 | Setae 0.8–1.5 mm; capsules weakly exserted, ellipsoid with a distinct and often somewhat inflated neck; operculum not clearly differentiated or weakly differentiated and non-functional; peristome absent | Bryobeckettia |
| 4' | Setae longer; capsules clearly exserted, not ellipsoid; operculum well differentiated and functional; peristome absent (in Physcomitrium) or more often well developed | 5 |
| 5 | Annulus strongly differentiated, compound, revoluble, composed of 1–4 rows of vesicular cells; exostome teeth fused at their apices by a lattice disc; capsules strongly sulcate at maturity; exothecial cells in alternating bands of thin- and thick-walled cells | Funaria |
| 5' | Annulus absent or rudimentary (in N.Z. species); exostome teeth either absent or not fused apically; capsules not sulcate; exothecial cells not in alternating bands of thin- and thick-walled cells | 6 |
| 6 | Peristome absent; calyptra mitrate; exothecial cell walls uniformly thickened when viewed in cross-section; spores spinose | Physcomitrium (pyriforme) |
| 6' | Peristome present or sometimes absent; calyptra mostly cucullate (mitrate in E. laxus, but then a double peristome present); exothecial cell walls cuneate when viewed in cross-section (but uniformly thickened in E. laxus); spores variously ornamented | Entosthodon |
Biostatus
Indigenous (Non-endemic)
| Category | Number |
|---|---|
| Indigenous (Endemic) | 1 |
| Indigenous (Non-endemic) | 10 |
| Exotic: Fully Naturalised | 1 |
| Total | 12 |
Bibliography
Fife, A.J. 1985: A generic revision of the Funariaceae (Bryophyta: Musci) Part I. Journal of the Hattori Botanical Laboratory 58: 149–196.
Fife, A.J. 2019: Funariaceae. In: Smissen, R.; Wilton, A.D. (ed.) Flora of New Zealand – Mosses. Fascicle 45. Manaaki Whenua Press, Lincoln.
Fife, A.J.; Seppelt, R.D. 2001: A revision of the family Funariaceae (Musci) in Australia. Hikobia 13: 473–490.
Goffinet, B.; Buck, W.R. 2011: Physcomitridium readeri is the correct name for Ephemerella readeri. Bryologist 114: 545–546.
Goffinet, B.; Buck, W.R.; Shaw, A.J. 2009: Morphology, anatomy, and classification of the Bryophyta. In: Goffinet, B.; Shaw, A.J. (ed.) Bryophyte Biology. Edition 2. Cambridge University Press, Cambridge. 55–138.
Hooper, E.J.; Duckett, J.G.; Cuming, A.C.; Kunin, W.E.; Pressel, S. 2010: Ephemerella readeri Müll. Hal. (Physcomitrella readeri (Müll. Hal.) I.G. Stone & G.A.M. Scott, Funariidae, Bryophyta): a genus and species new to Europe. Journal of Bryology 32: 256–264.
Lang, D.; Zimmer, A.D.; Rensing, S.A.; Reski, R. 2008: Exploring plant biodiversity: the Physcomitrella genome and beyond. Trends in Plant Science 13(10): 542–549.
Liu, Y.; Budke, J.M.; Goffinet, B. 2012: Phylogenetic inference rejects sporophyte based classification of the Funariaceae (Bryophyta): Rapid radiation suggests rampant homoplasy in sporophyte evolution. Molecular Phylogenetics and Evolution 62: 130–145.
McDaniel, S.F.; von Stackelberg, M.; Richardt, S.; Quatrano, R.S.; Reski, R.; Rensing, S.A. 2010: The speciation history of the Physcomitrium—Physcomitrella species complex. Evolution 64: 217–231.
Rensing, S.A.; Lang, D.; Zimmer, A.D.; Terry, A.; Salamov, A.,; Shapiro, H.; Nishiyama, T.; Perroud, P.-F.; Lindquist, E.A.; Kamisugi, Y.; Tanahashi, T.; Sakakibara, K.; Fujita, T.; Oishi, K.; Shin-I, T.; Kuroki, Y.; Toyoda, A.; Suzuki, Y.; Hashimoto, S.; Yamaguchi, K.; Sugano S.; Kohara, Y.; Fujiyama, A.; Anterola, A.; Aoki, S.; Ashton, N.; Barbazuk, W.B.; Barker, E.; Bennetzen, J.L.; Blankenship, R.; Cho, S.H.; Dutcher, S.K.; Estelle, M.; Fawcett, J.A.; Gundlach, H.; Hanada, K.; Heyl, A.; Hicks, K.A.; Hughes, J.; Lohr, M.; Mayer, K.; Melkozernov, A.; Murata, T.; Nelson, D.R.; Pils, B.; Prigge, M.; Reiss, B.; Renner, T.; Rombauts, S.; Rushton, P.J.; Sanderfoot, A.; Schween, G.; Shiu, S.-H.; Stueber, K.; Theodoulou, F.L.; Tu, H.; van de Peer, Y.; Verrier, P.J.; Waters, E.; Wood, A.; Yang, L.; Cove, D.; Cuming, A.C.; Hasebe, M.; Lucas, S.; Mishler, B.D.; Reski, R.; Grigoriev, I.V.; Quatrano, R.S.; Boore, J.L. 2008: The Physcomitrella genome reveals evolutionary insights into the conquest of land by plants. Science, an Illustrated Journal Published Weekly 319(5859): 64–69.
Schwägrichen, C.F. 1830: Species Muscorum Frondosorum. Vol. 5. Part 1. Nauck, Berlin.
Wettstein, F. von 1932: Genetik. In: Verdoorn, F. (ed.) Manual of Bryology. Nijhoff, The Hague. 233–272.