Class
Family
The following generic description is modified from Frahm (1994a).
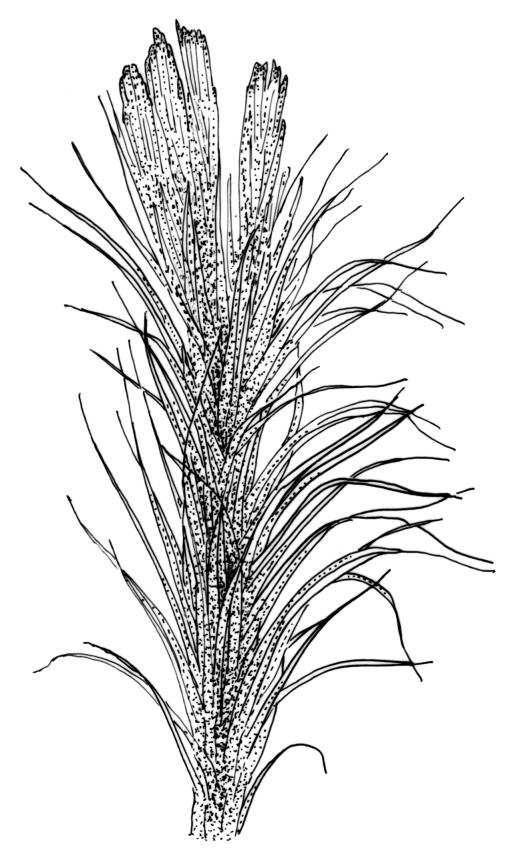
Plants erect in loose to compact tufts or turves, variable in height, mostly uniformly foliate, rarely comose, sometimes fragmenting. Stems simple or seldom forked, in cross-section with a central strand, densely or sparsely beset with rhizoids. Leaves mostly lanceolate, lacking a distinct shoulder, mostly acuminate or acute, occasionally rounded or cucullate, erect or curved, straight or ± flexuose; upper laminal cells quadrate, short-rectangular, weakly elongate, or rhombic; cells of the leaf base enlarged, with thin or thick walls, sometimes porose, sometimes elongate at margins; alar cells absent or conspicuously differentiated, hyaline or red-brown, often extending to the costa. Costa broad, usually more than ½ the width of the leaf base, distinctly or weakly differentiated from the lamina, lacking oil-inclusions, percurrent or excurrent, often ending in a hyaline apex, in mid leaf cross-section with 0, 1, or 2 stereid bands, often ribbed and sometimes toothed on the abaxial surface, with or without enlarged cells (hyalocysts) on the adaxial surface.
Dioicous. Perichaetia terminal but often overtopped by innovations, with perichaetial leaves broader than vegetative leaves, sheathing in N.Z. species. Setae elongate, nearly always cygneous; capsules erect or inclined, ovoid to short-cylindric, strumose or not, often furrowed, especially when dry; annulus well differentiated, vesicular and revoluble in N.Z. species; stomata absent; operculum rostrate. Peristome teeth 16, deeply divided, vertically pitted-striolate below. Calyptra cucullate, smooth, usually fringed at base. Spores mostly less than 20 µm, smooth.
Campylopus is a large genus found in all parts of the world except polar regions, with most species growing on acidic or nutrient-depleted soil. It is most diverse in the Americas. It is taxonomically a very difficult genus in all parts of its range due to its morphological plasticity; Crum & Anderson (1981) characterised it as "one of the most difficult genera of mosses". Campylopus and some closely allied genera are sometimes isolated within the Dicranaceae into a subfamily Campylopodioideae (Frahm 1985) on the basis of their cygneous setae, lack of stomata, broad costae, and frequent presence of enlarged cells on the adaxial surface of the costa. Some authors (Goffinet et al. 2009) have placed Campylopus in the Leucobryaceae, but this is not done here.
Little reliance can be given to the few published estimates of the number of species in the genus. For a very rough scale, Brotherus (1924) estimated it might contain about 500 species worldwide, while an even larger figure of c. 750 species is given by Corley (in Smith 2004). Frahm (1994b) accepted 25 species from Australia, but his species concepts seem excessively narrow. N.Z. material, like that from many parts of the generic range, can be extremely difficult to name with confidence even when broad species concepts are applied.
Campylopus is mostly readily recognisable by its terrestrial habitat and erect habit with very broad costae (more than ½ the width of the basal part of the leaf) and often conspicuously differentiated alar cells. It rarely fruits, but capsules when present have cygneous setae and calyptrae that are usually fringed at the base. Stomata are universally absent in the genus.
The genus is traditionally divided into three subgenera (Brotherus 1924) on the basis of costal anatomy, which is best seen in cross-sections made at mid leaf. Some recent regional treatments do not make use of these subgeneric distinctions (Frahm 1994b; 1994a; Allen 1994) while recent treatments for eastern North America (Crum & Anderson 1981) and the British Isles (Corley in Smith 2004) assign species to subgenera. Regardless, all treatments require observations on costal anatomy for specimen identification.
Despite the modest number (six) of species accepted here, their great plasticity and the rarity of capsule production frequently make confident identifications difficult. Some collections will remain unnameable. Sadly, but inevitably, these unnameable collections include some type specimens.
Some N.Z. authors have liberally described taxa on the basis of arguably aberrant collections of known species. Brown (1897), for example, described 15 N.Z. species in one paper; the types of only a few are available in N.Z. herbaria. Carl Müller (especially in Müller & Brotherus 1900) likewise described many N.Z. Campylopus taxa, mostly based on material collected by H. Schauinsland. Many of Brown’s names were treated as synonyms by Dixon (1923) and/or by Robinson (1975). Many of the types of these names are not accessible, and Dixon’s and Robinson’s synonymies are not questioned here. Robinson placed eight Robert Brown (bis) names in the synonymy of C. clavatus and four others in the synonymy of C. appressifolius (itself considered here a synonym of C. clavatus). Brown’s inclination to describe trivial variants as new species is well documented (Dixon (1913). Müller, particularly in the latter part of his career, was also known for basing new species on minor variants of described taxa. In the single 1900 paper he described six new Campylopus species based on N.Z. (including Chatham I.) material.
The German bryologist J.-P. Frahm published numerous papers on Campylopus in Australasia and other regions between 1981 and 1994. In a N.Z. context, he described several taxa that are not recognised here, and recorded taxa that were originally described from outside Australasia (e.g., Campylopus catarractilis). Some of these records are not accepted in this treatment. Although the illustrations in Bartlett & Frahm (1983) are useful in the recognition of N.Z. species, the often contradictory opinions expressed in Frahm’s several publications limit their value in the interpretation of our taxa.
This treatment employs broader species concepts than have been applied to N.Z. Campylopus by previous authors. It is presented here, warts and all, as a "state-of-current-knowledge" treatment rather than a definitive one.
In the key and the descriptions that follow, all costal sections are taken at mid leaf unless specifically stated otherwise. The phrase "upper laminal cells" likewise refers to cells at mid leaf. Leaf sections are best taken by isolating 2–3 representative leaves in a small water drop and cutting numerous sections using a single-edge razor blade. This can be done under a stereoscope while pinning the leaf to the slide, convex side up, with a pair of jeweller’s forceps. If basal costal sections are required, these can usually be cut from the same leaf, at the approximate upper limit of the alar cells, before a cover slip is applied. The basal costal sections can nearly always be readily differentiated by their size and outline from the mid costal sections.
| 1 | Apices of some or all vegetative leaves cucullate | 2 |
| 1' | Apices of vegetative leaves not cucullate (cucullate leaves may occur on propagula, as in C. purpureocaulis) | 3 |
| 2 | Shoots often serially comose; costa usually with short lateral spurs, not filling the upper subula, in mid leaf cross-section lacking adaxial hyalocysts, with a few small and thick-walled adaxial cells and median guide cells; basal laminal cells thick-walled and porose to insertion or very nearly so | Campylopus kirkii |
| 2' | Shoots not serially comose; costa lacking lateral spurs, often filling the upper subula, in mid leaf cross-section with exposed adaxial hyalocysts; basal laminal cells forming a distinct group of thin-walled and non-porose cells near insertion | Campylopus bicolor |
| 3 | Costa in cross-section (mid leaf) with stereids exposed on adaxial surface | 4 |
| 3' | Costa in cross-section (mid leaf) with hyalocysts exposed on adaxial surface | 5 |
| 4 | Propagula (apical plantlets) distichous; upper laminal cells (at mid leaf) small and irregular, at least some subquadrate, incrassate, not porose; costal cross-section (mid leaf) with well developed stereid bands on both sides of a median layer of guide cells, the adaxial stereid layer of 2–3 or more cell layers; capsules ovoid, curved and sulcate when dry, smooth at base; peristome teeth vertically striate below, split c. ⅔ their length | Campylopus purpureocaulis |
| 4' | Propagula (apical plantlets) stellate; upper laminal cells (at mid leaf and upwards) mostly elongate-rhombic or rounded-rectangular, sometimes irregularly elongate, weakly to strongly porose, ± obliquely oriented; costal cross-section (mid leaf) with a single layer of exposed adaxial stereids; capsules subcylindric, suberect, and smooth or nearly smooth when dry, scabrid at base; peristome teeth coarsely and irregularly lirate throughout, split nearly to base | Campylopus clavatus |
| 5 | Leaves of upper shoots mostly ending in a squarrose, hyaline or yellow hair point; plants lacking deciduous falcate leaves, comose (often serially) when fertile; upper laminal cells mostly rhombic and ± rounded at corners, obliquely oriented; costa in cross section (mid leaf) with 1-celled abaxial ridges (rarely merely crenulate); rhizoids ± brown…. | Campylopus introflexus |
| 5' | Leaves of upper shoots not squarrose, concolourous at apex or with a short hyaline tip; plants often with masses of pale and strongly falcate deciduous leaves, not comose when fertile; upper laminal cells mostly quadrate or short-rectangular, not obliquely oriented; costa in cross-section (mid leaf) lacking abaxial ridges; rhizoids brick-red | Campylopus pallidus |
| Category | Number |
|---|---|
| Indigenous (Non-endemic) | 6 |
| Total | 6 |
Campylopus and Thysanomitrium
C. acuminatus Mitt. [J. Linn. Soc., Bot. 12: 90 (1869)] has been recorded from N.Z. by Frahm (1994b, and elsewhere) from Hermite I. is not accepted here as a N.Z. plant for reasons discussed under C. kirkii.
Campylopus capillaceus Hook.f. & Wilson [London J. Bot. 3: 543 (1844)] is an earlier name for what has been termed Campylopodium medium (Duby) J.-P.Frahm & Giese, but which is treated in this eFlora as Campylopodium capillaceum (Hook.f. & Wilson) Fife, comb. nov. The name Campylopus capillaceus sensu Fife 1995 is a misapplication of C. capillaceus Hook.f. & Wilson, and the more appropriate name for such material is C. pallidus, q.v.
Campylopus catarractilis (Müll.Hal.) Paris [Index Bryol. Suppl. 90 (1900)] is a South African species recorded from the Poor Knights Is (Beever 1986; Beever et al. 1992), based on determinations by J.-P. Frahm. However, re-examination of the relevant material shows that this material is a form of C. introflexus, q.v.
Campylopus distractus Müll.Hal. in Müll.Hal. & Broth. [Abh. Naturwiss. Vereins Bremen 16(3): 495 (1900)] Type: N.Z., W. Walker (Herb. Levier). There is no material in H-Br. fide Frahm (1998, p. 31). Dixon (1923, p. 85) indicated that he saw the type of "Dicranum distractum Müll.Hal." [Hedwigia 36: 350 (1897)]. This type was collected by W. Walker, s. loc., in 1875, and cited by Müller as being in the Levier herbarium. Müller’s name was transferred to Campylopus by Paris (1900, p. 91). It is not discussed further here.
Campylopus flavoviridis Dusén [Ark. Bot. 4(13): 2 (1905)], see C. incrassatus, below.
Campylopus holomitrium sensu Sainsbury 1955. See discussion under C. pallidus.
Campylopus incrassatus Müll.Hal. [Linnaea 18: 686 (1845)]. This South American species is recorded from N.Z. by Frahm (1987), who cited an earlier report of C. flavoviridis Dusén by Bartlett & Frahm (1983). I have seen two collections from the North I. named as C. incrassatus by J.-P. Frahm; both are clearly referable to C. pallidus. No material named as C. flavoviridis appears to be present in any N.Z. herbarium. Neither C. incrassatus nor C. flavoviridis is accepted here as N.Z. species.
Campylopus kirkii var. pilosus J.-P.Frahm [Lindbergia 7: 30, 1981] is rejected here for reasons discussed under C. kirkii.
Campylopus leptodus Mont. [Ann. Sci. Nat., Bot., sér. 3, 4: 111 (1845)] was recorded (as a Thysanomitrium) and discussed by Dixon (1923, p. 91). The original material of Campylopus leptodus is Chilean. Dixon apparently saw material from "principally near Auckland" and collected by both Colenso and Knight. He considered it to differ from nearly all other N.Z. Campylopus species, with the notable exception of C. purpureocaulis, by having both ventral and dorsal stereid bands (Palinocrapsis costal anatomy). Dixon differentiated the Colenso and Knight material from C. purpureocaulis (cited as C. arboricola) by having "larger, longer leaves which (except the comal, floral ones) do not end in the distinct white hair-point which is a marked character of that plant", as well as its large inflated alar cells.
Some collections referable to C. purpureocaulis (e.g., G.O.K. Sainsbury 1999 from Mt Taranaki, CHR 566304) have predominantly concolorous, non-hair-pointed leaf apices. Also, C. purpureocaulis usually has well-differentiated, inflated, and pigmented alar cells, although these are often obscured by the abundant rhizoids. Dixon’s observations on leaves of the Colenso and Knight material suggest that it represented slightly aberrant C. purpureocaulis. His suggestions that the material, when fruiting, had a "longly fringed calyptra and [a] narrow, smooth, symmetrical capsule" is more suggestive of C. clavatus, a species with a similar costal structure, and one often placed in the genus (or subgenus) Thysanomitrium. Material named as C. leptodus in the Beckett collection belongs to the unrelated Saelania glaucescens and is not pertinent. Campylopus leptodus has not, to my knowledge, been considered in any work on Australasian Campylopus since that of Dixon. On the balance of the limited available evidence it is likely that the specimens cited by Dixon (1923) as Thysanomitrium leptodus are referable to either C. purpureocaulis or C. clavatus. Neither C. leptodus nor Thysanomitrium leptodus are considered further in this treatment.
Campylopus paludosus J.-P.Frahm & J.K.Bartlett in J.K.Bartlett & J.-P.Frahm [J. Bryol. 12: 376 (1983)]. Although I am unable to refer type material from near Jacksons [Westland L.D., J.K. Bartlett 63, AK 221722] to any N.Z. species known to me, I am reluctant to conclude, as did Bartlett & Frahm, that this collection deserved description as a new species. C. paludosus is here considered as nom. dub. and not discussed further in this eFlora. Bartlett’s illustration (Bartlett & Frahm, 1983, fig. 8) is an accurate representation of the Jacksons collection.
Campylopus pyriformis var. hamatus J.-P.Frahm & J.K.Bartlett in J.K.Bartlett & J.-P.Frahm [J. Bryol. 12: 378, 1983]. The type collection is from the Denniston Plateau (Nelson L.D., J.K. Bartlett 59, Holotype: WELT M007126; Isotype: AK 221721). I concur with Bartlett & Frahm (1983) that the nature of upper laminal cells and the costal cross-section indicate a probable relationship to C. pallidus (C. pyriformis). However, I am reluctant to conclude that this collection deserves recognition as a variety of the parent species and it is likely just aberrant material of that species. The var. hamatus is not discussed further in this eFlora. Bartlett’s illustration (Bartlett & Frahm 1983, fig. 10) is an accurate representation of the type.
Campylopus vesticaulis Mitt. in Melliss [St. Helena 359 (1875)] was described from the island of St. Helena; it is not accepted as part of the NZ flora. This name was apparently first used in a N.Z context by Beever et al. (1992), on advice of J.-P. Frahm, to replace C. paludosus. Campylopus vesticaulis was included in Fife (1995). Frahm did not apply this name consistently to N.Z. material and the occurrence of this species in N.Z. is deemed phytogeographically unlikely. The only specimen seen, apart from the type of C. paludosus (see above), named by Frahm as C. vesticaulis (Frahm 22-15 from Mt Ruapehu, CHR 504783) is referable to C. clavatus. It is not discussed further here.
Campylopus xanthophyllus Mont. [Ann. Sci. Nat., Bot., sér. 3, 4: 111, 1845)] is a South American species recorded from two N.Z. localities by Wilson (1854, p. 68). Dixon (1923, p. 91) opined that the species was not adequately documented here and that it should be excluded from our flora. It is not discussed further here.