Class
Family
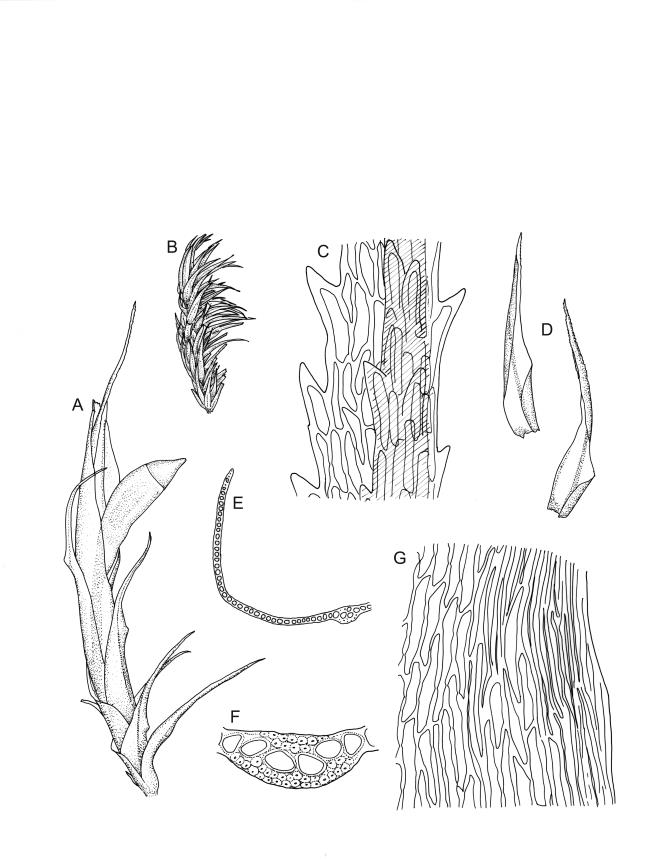
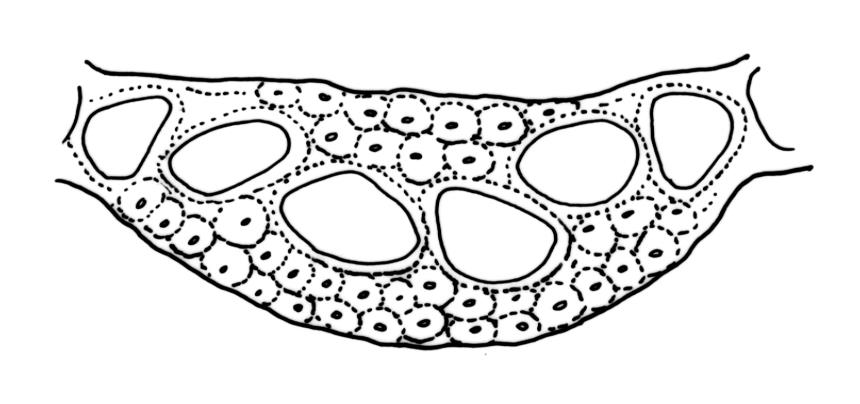
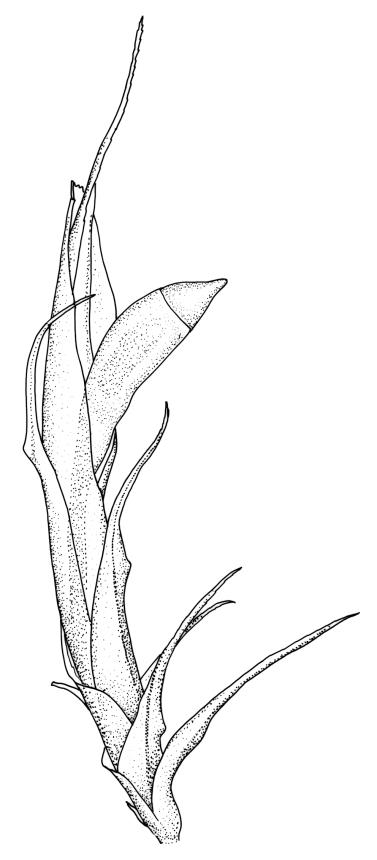
Plants medium-sized, white-green when fresh, becoming gold-brown in dried specimens, forming cushions. Stems 20–35 mm, much branched by forking and innovation, often bearing multiple terminal perichaetia, in cross-section with a weak and ill-defined central strand, and 3–5 layers of thick-walled cortical cells; densely matted throughout by short, tangled, smooth, and pale-brown or nearly white rhizoids. Leaves secund, curved but not falcate, variably plicate when fresh, not rugose, ovate-lanceolate, scarcely narrowed at insertion, sharply spinose-serrate in upper ⅓ or less, 4.0–5.5(–6.5) × 0.8–1.0 mm (under cover slip), subtubulose, plane at margins; mid laminal cells elongate, ± irregular, 80–120 × 7–10(–13) µm and 7–12:1, thick-walled and porose, with elongate cells extending to apex; juxtacostal cells at mid leaf not differentiated; border well defined and broad, pale, extending from just above the alar group to base of the serrations, 6–8 cells and c. 30–45(–75) µm wide at mid leaf; cells of leaf base slightly longer but otherwise as those of mid lamina; alar cells strongly and abruptly differentiated, unistratose, extending c. ½ to costal base and c. 6–8 cells up the margin, inflated and yellow, firm-walled, the largest c. 75 × 40 µm. Costa narrow, subpercurrent or percurrent, usually extending basipetally to the insertion and c. 60 µm and c. 1/20 or less the width of the leaf base, rarely vanishing in lowest ⅓ of leaf, the abaxial surface in upper c. ¼ bearing short spines in 2 ranks (scarcely visible under hand-lens), in mid leaf cross-section not winged, c. 5 cell-layers thick, with a median layer of 4–6 guide cells and one abaxial and one adaxial stereid group, which extend the width of the costa.
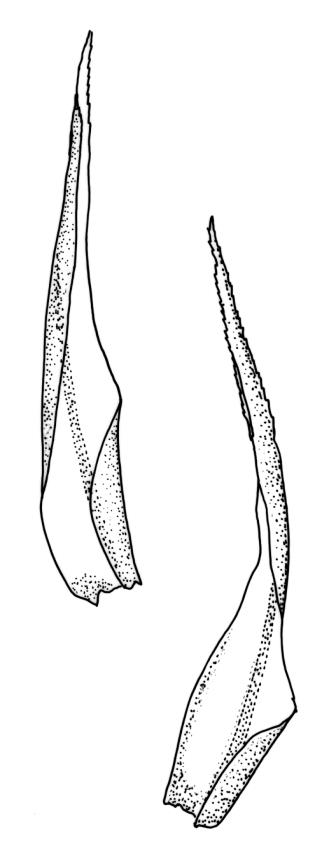
Pseudautoicous (or occasionally with male not dwarfed). Perichaetial leaves sheathing, strongly tubulose, and with costa extremely weak or lacking, the outer leaves abruptly narrowed from a tubulose base to long arista, the innermost leaves much longer, c. 12 mm, narrowly lanceolate and gradually tapered to a slender and toothed acumen, extending to or conspicuously exceeding the mouth of mature capsules. Dwarf males usually gemmiform, to c. 0.7 mm long, embedded in rhizoids of sterile or ♀ plants. Setae 1–3(–4) per perichaetium, 8–10 mm, straight, not twisted, pale brown; capsules emergent from one side of the perichaetium, curved, cylindric, 1.8–3.0 mm, weakly strumose, smooth when moist, constricted below the mouth and weakly sulcate when dry; exothecial cells firm-walled and ± oblong-hexagonal; stomata restricted to neck, superficial; annulus weakly differentiated, not revoluble; operculum curved-rostrate from a conic base (poorly illustrated here), ± equal the length of the capsule. Peristome teeth inserted at mouth, red-brown, split ½– ⅗ to base into segments of unequal width, 475–580 × 90–115 µm, papillose-striolate in lower c. ⅔, baculate near apex. Calyptra cucullate, smooth. Spores spherical, 15–18 µm, smooth.
Beckett 1894, pl. 30A (as Dicnemon obsoletinerve); Klazenga 2003, fig. 15; Malcolm & Malcolm 2006, pp. 72, 272, 295, 306.
The leaf plications in D. fasciatum are mostly weak but variable. They are most distinct in dry material and in some collections are suggestive of the weak plications that occur in D. plurisetum, the species with which it is most likely to be confused. The very strong leaf border (limbidium) of D. fasciatum is comparable in width to that in D. plurisetum. However, D. fasciatum differs from D. plurisetum by shorter (4–6.5 vs. 8–10 mm) vegetative leaves and by emergent rather than exserted capsules. Dicranoloma fasciatum also differs markedly from D. plurisetum by the shape of its perichaetial leaves.
NI: N Auckland, including offshore islands (GB, LB, RT), S Auckland; Gisborne (Manganuku Stream, Hikurangi, Lake Waikaremoana), Hawke’s Bay (Pētane, Hutchinson Domain Scenic Reserve), Taranaki (Tāngarākau Gorge, Pūniho Track, Te Maire Scenic Reserve), Wellington (Ōhakune, Tangiwai, Tararua Range); SI: Nelson, Marlborough, Canterbury, Westland, Otago, Southland (Manapōuri, Bluff); St. The report from Auckland Is by Vitt (1979) is based on misidentifications of D. robustum.
Endemic or Australasian. Klazenga (2003) tentatively recorded this species from a small number of scattered localities in N.S.W., Vic., and King I. These records were based on non-fruiting material that he tentatively placed here. He considered this material to "belong to D. fasciatum because the leaf length is the same as in New Zealand specimens of D. fasciatum" but treated the determinations as "not definitive".
On rotten logs, tree bases, exposed roots, and lower trunks (e.g., of Fuscospora fusca, Griselinia littoralis); less commonly on duff or soil. This species is often epiphytic on tree ferns and on rotting bases of dead asteliads. Occurring in a variety of forest types, including broadleaved-podocarp and Leptospermum scoparium-dominated forests, as well as southern beech forest. Commonly associated with D. menziesii (less often with other species of Dicranoloma), Leucobryum javense, Rhizogonium distichum, Wijkia extenuata, and the hepatics Bazzania adnexa, Heteroscyphus coalitus, and Cuspidatula monodon. Ranging from c. 3 m (Little Barrier I., North Auckland L.D.; J.E. Beever, pers. comm., Sept. 2017) to at least 720 m (Bells Junction, Wellington L.D.) on the North I. and its associated islands, and from near sea level (at several localities) to 960 m (Cobb Valley, Nelson L.D.) on the South I.
This is a relatively uncommon species, although it is particularly well documented from localities in N Auckland, S Auckland, and Banks Peninsula. Sterile material cannot always be determined with confidence. When fertile the elongate perichaetial leaves, which extend to or well beyond the capsule mouth, are its most distinctive feature. In the field the plants are a white-green colour, similar to that in D. dicarpum, but they turn gold-brown in dry material. D. fasciatum forms cushions rather than the extensive turves formed by many of its congeners. Only D. menziesii has perichaetial leaves of comparable length relative to the sporophytes. Dicranoloma fasciatum is a much paler plant than D. menziesii, and it also differs by having much shorter leaves and elongate upper laminal cells.
Material of D. fasciatum from Hari Hari (Westland L.D., L. Visch 510, CHR 267056) is unusual because it has non-dwarf male plants. The ♂ plants have stems to c. 8 mm (roughly half the height of the associated ♀ stems), vegetative leaves representative of the species, and numerous lateral and terminal perigonial branches of less than 1 mm length. Each gemmiform perigonium has yellow, ovate-lanceolate bracts to 1.8 mm, numerous antheridia, and filiform, 7–8-celled paraphyses. Similar ♂ plants, approximately ⅔ the height of associated ♀ plants, have been seen from Southland (K.W. Allison 3484, CHR 542638).
Populations in which the costae are weak or vanish completely in the lower portion of the leaf (B. Wormald 27 from Tararua Range, Wellington L.D., CHR 524330) are comparatively rare, and this feature is given undue emphasis by Sainsbury (1955, p. 135) in his description.
Hedwig (1801, p. 127) cited the type locality of Dicranum fasciatum as "Insulae maris pacifici", and this is presumed to be N.Z. It is unclear who might have collected it prior to 1801, although A. Menzies is a possibility. Correspondence in 2000 with the then curator of the Hedwigian herbarium at Geneva, Phillipe Clerc (pers. comm., 8 June 2000), failed to locate any potential type material. Klazenga (2003, p. 447) also failed to locate type material and lectotypified Dicranum fasciatum Hedw. using Spec. Musc. Frond. Plate 28. Hedwig’s illustration is a convincing rendition of the N.Z. plant, which was termed Dicranoloma fasciatum (Hedw.) Paris by both Dixon (1913, p. 26) and Sainsbury (1955). There is no evidence that Dixon examined Hedwig’s type of Dicranum fasciatum.
Dixon’s (1913, p. 26) judgement that Dicnemon obsoletinerve Hampe & Müll.Hal. is synonymous with Dicranum fasciatum Hedw. was accepted by Klazenga (2003) and is accepted here. Klazenga expressed doubt concerning the collector, and by implication the collection site, of the material he selected as the lectotype. Material from Kaipara, N Auckland L.D. (S. Mossman 712, NY-Mitten) in 1850 and apparently determined by C. Müller as Dicranum fasciatum, has been examined; given its collection locality and date it could be part of the original collection apparently sent to F. Mueller (see Klazenga 2003, p. 447). It is noteworthy that C. Müller described other species of Dicranoloma (e.g., Dicranum brachypelma) from Mossman collections.
Other collections of Dicnemon obsoletinerve Hampe cited by Dixon were most probably collected by T.W.N Beckett at either Fishers Bush, near Oxford, Canterbury L.D., in March 1884 (CHR 542796) or at Studholme Bush, near Waimate, Canterbury L.D., in September 1888 (CHR 543117). Both these collections are represented in the Beckett herbarium, and portions of both were identified by C. Müller as Dicnemon obsoletinerve.