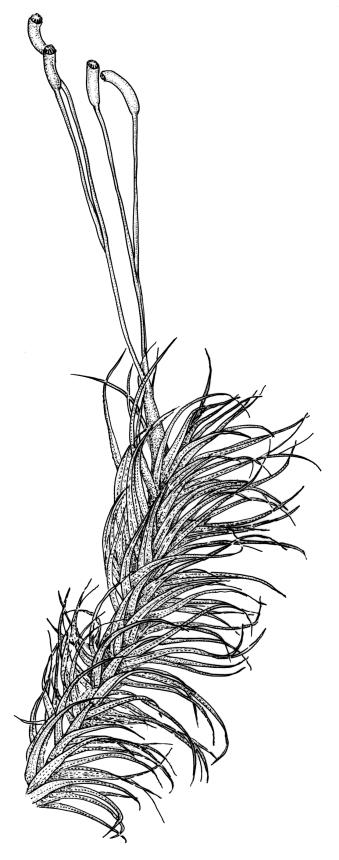
Class
Family
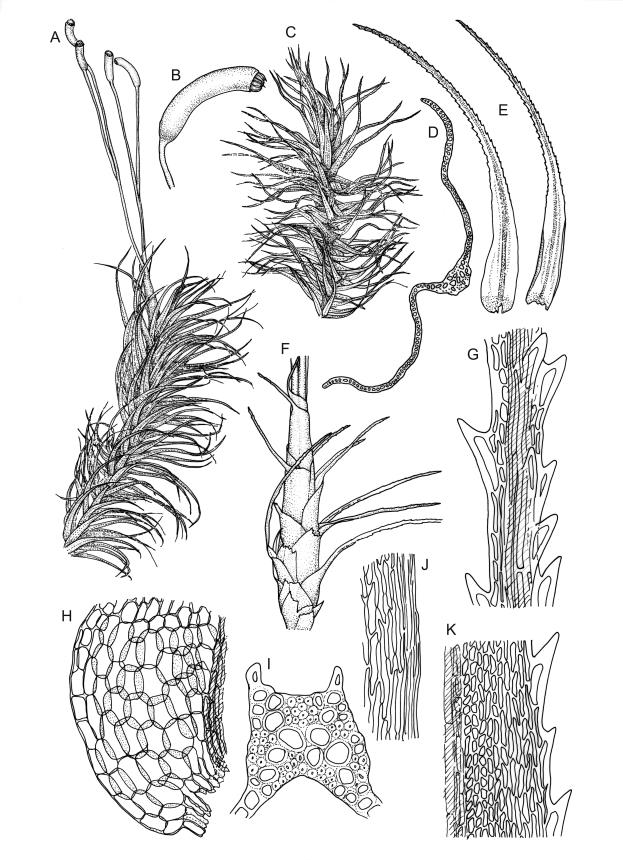
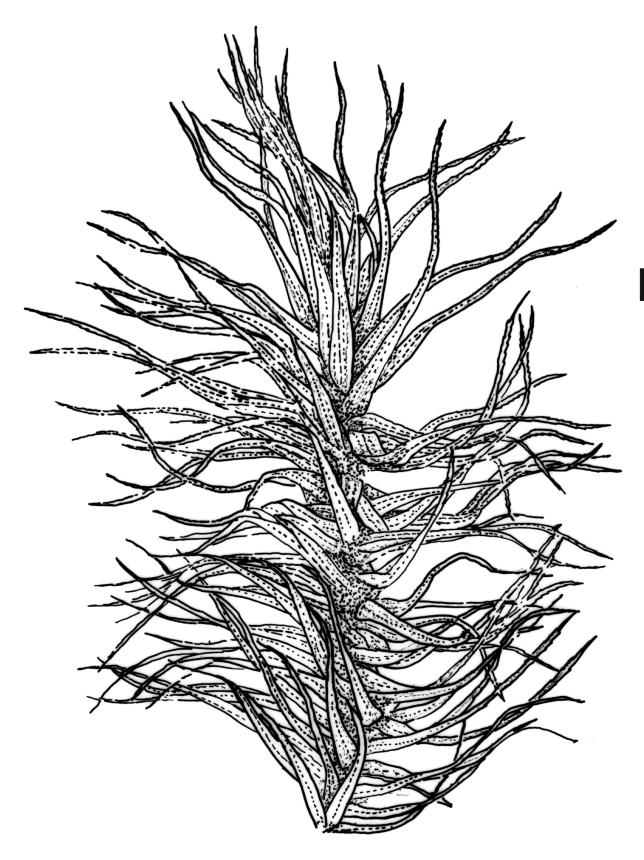
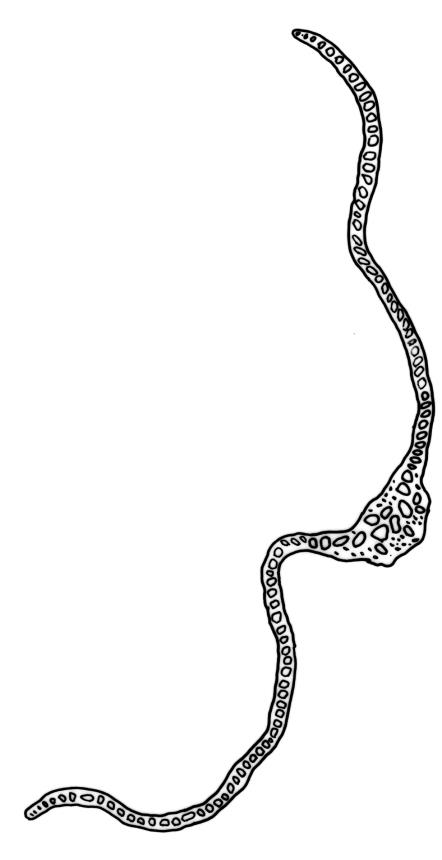
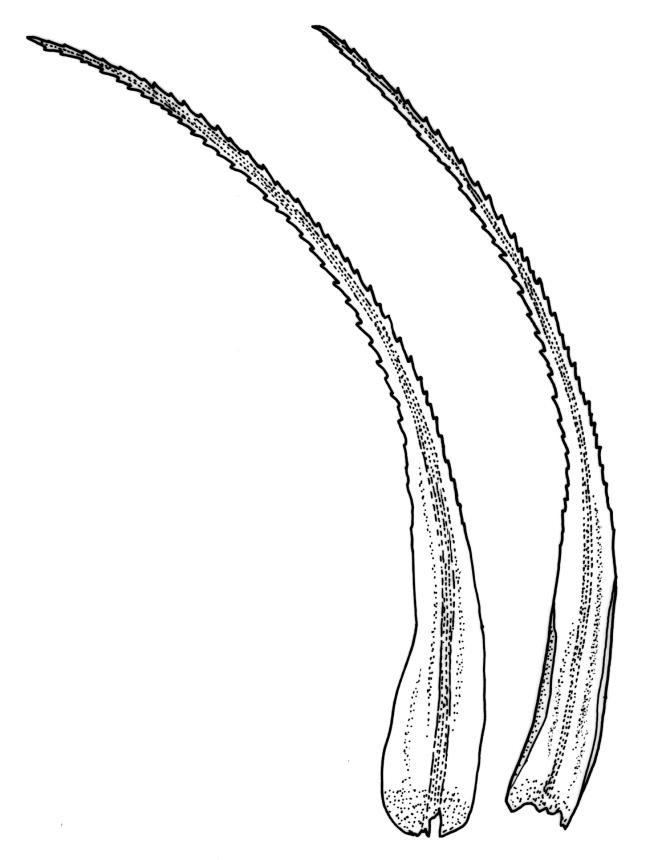
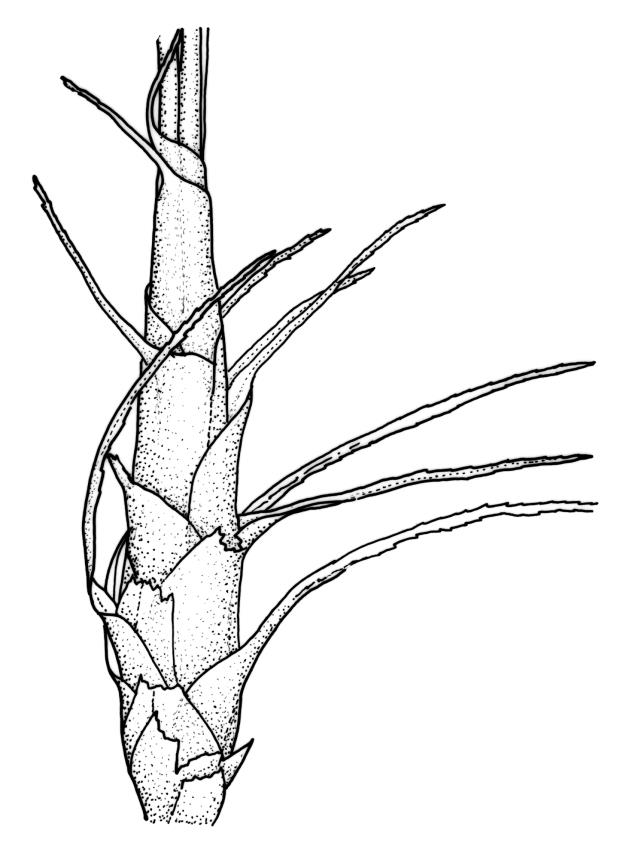
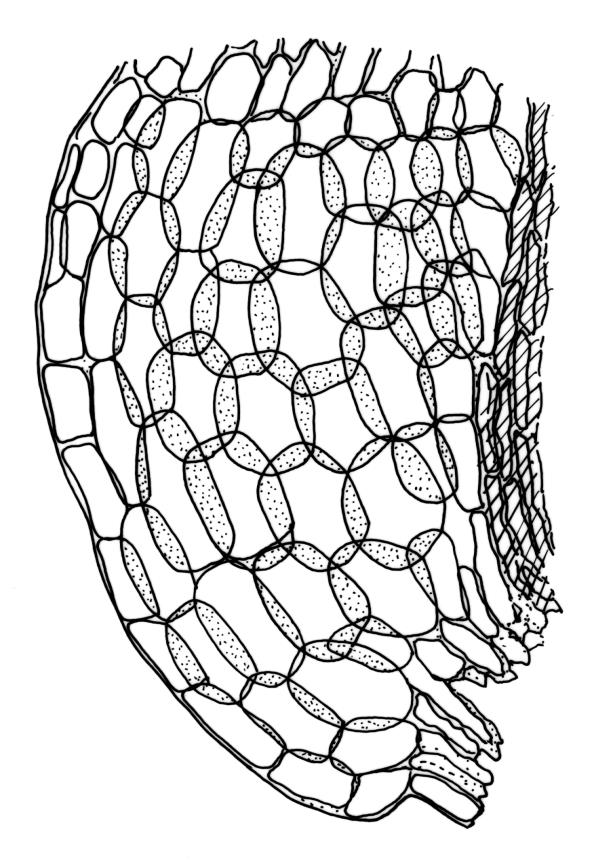
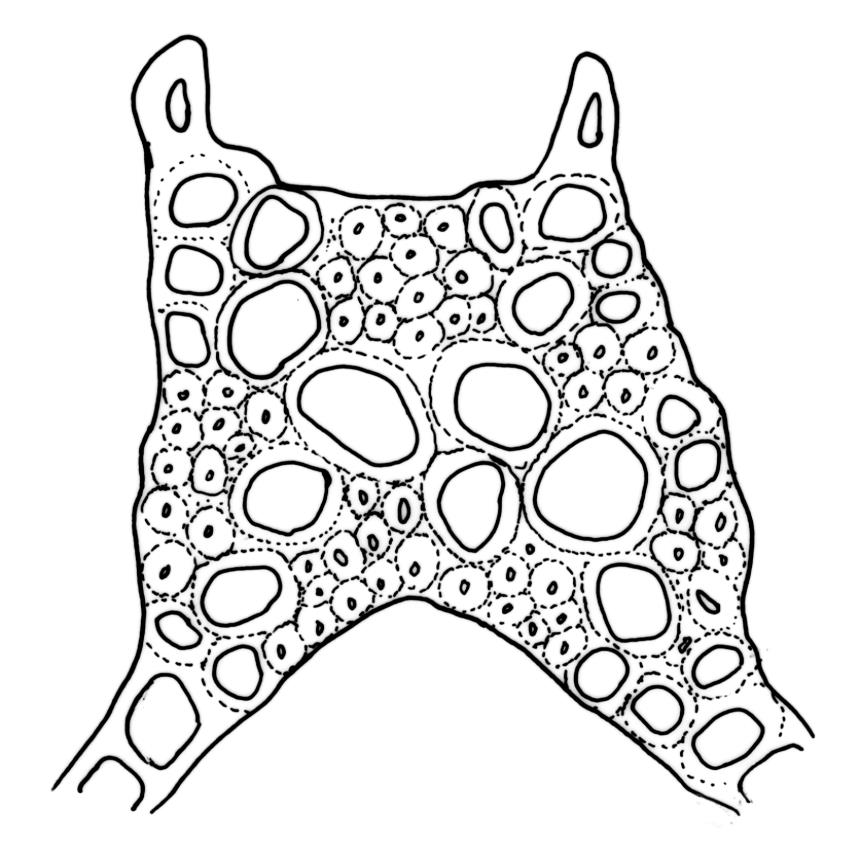
Plants robust, white- or pale yellow-green when fresh, becoming gold-brown in older specimens, forming tufts, cushions, or turves. Stems c. 35–70(–90) mm, little or sparsely forked, in cross-section with 2–4 layers of thick-walled cortical cells and an ill-defined central strand, densely matted throughout by short, tangled, smooth, and pale brown or whitish rhizoids. Leaves mostly strongly secund from a spreading base, ± falcate, strongly plicate, somewhat more flexuose but otherwise little altered when dry, narrowly lanceolate, sharply spinose-serrate in upper ½–⅔, (6–)7–10 × c. 0.7–1.0 mm, keeled, plane at margins; mid laminal cells elongate, mostly c. (45–)60–80(–100) × 6–10 µm, strongly pigmented, porose, outermost cells linear and pale; juxtacostal cells at mid leaf shorter and irregular in outline, mostly 1–4:1, more pigmented, and less porose than contiguous laminal cells; border narrow and very pale (especially in leaf base), extending from just above the alar group to the marginal teeth, mostly of 1–4 linear cells and c. 18–30 µm wide at mid leaf; cells of leaf base more elongate than those at mid leaf, mostly 120–180 × 6–12 µm, thick-walled and porose; alar cells strongly and abruptly differentiated, unistratose, extending c. ⅔ or more to costal base and c. (6–)8–10 cells up the margin, inflated and yellow, firm-walled, the largest c. 90 × 60 µm. Costa occupying c. ⅛ the widest part of leaf base, nearly filling the upper ⅓ of leaf, the abaxial surface in upper ½ or more bearing single-celled spines (obvious under hand-lens), in cross-section strongly 2-winged at mid leaf, with 7–10 median guide cells (with 4 often forming a central group), abaxial and adaxial stereid groups, and 2–4 ± isolated and larger cells on the abaxial surface.
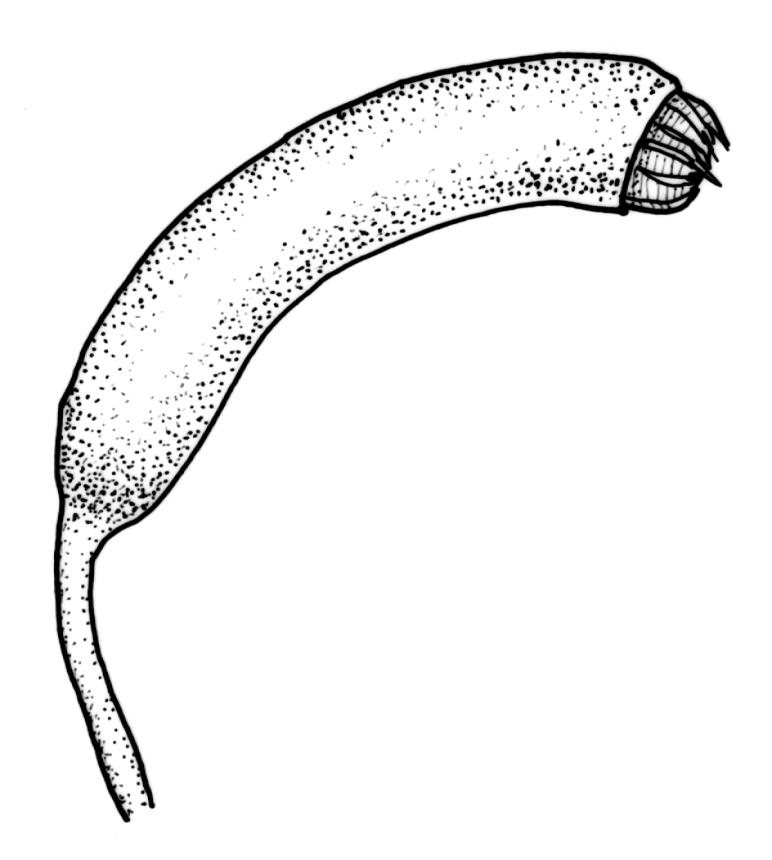
Pseudautoicous. Perichaetial leaves sheathing (up to c. ¼ the seta length) and strongly concave, abruptly tapered from an obovate or elliptic base to a long and slender subula. Dwarf males gemmiform, c. 0.5–0.6 mm long, usually embedded in rhizoids of a sterile or ♀ plant, occasionally epiphytic on vegetative leaves, with few (<3?) antheridia. Setae aggregated, 4–6(–9) per perichaetium, c. 18 mm, straight or weakly flexuose, scarcely twisted, pale brown; capsules exserted, strongly curved, cylindric, 3–3.5 mm, not or weakly strumose, smooth when moist, constricted below the mouth when dry; exothecial cells firm-walled and oblong-hexagonal; stomata restricted to neck, superficial; annulus a single row of weakly differentiated cells, not revoluble; operculum curved-rostrate from a conic base, 2–3 mm. Peristome teeth inserted at mouth, red-brown, split c. ½ to base into unequal segments, c. 450 × 80 µm, papillose-striolate nearly throughout. Calyptra cucullate, smooth. Spores ellipsoid to spherical, 18–21 µm, nearly smooth.
Dixon 1913, pl. 8, fig. 8 a–c; Brotherus 1924, fig. 166; Norris & Koponen 1990, fig. 12 a–d (as Dicranum dicarpum); Klazenga 2003, fig. 11.
In the field D. dicarpum is highly distinctive given its grey-green colour and strongly plicate leaves; the pale coloration is often lost in older herbarium material. The very slender upper leaf with narrow laminae and densely and sharply spinose-toothed margins and costal abaxial surface provide further distinction. Under the stereoscope the width of the leaf at the insertion (when still attached to the stem) is ≤1 mm, but this is often obscured by rhizoids. Under the compound microscope an obvious juxtacostal group of short, ± opaque, irregular, and scarcely porose cells usually begins c. 1–1.5 mm above the alar cells and extends far up the leaf. The juxtacostal group of short cells can be up to one half the width of the lamina at mid leaf, and can be seen under a hand-lens; it provides a reliable character differentiating this species from its N.Z. congeners.
Dicranoloma dicarpum is sometimes confused with D. plurisetum, which has a similar white- or grey-green coloration when fresh. D. dicarpum is a generally finer plant in appearance, with narrower leaves (c. 0.7–1.0 mm wide vs c. 1.5–1.8 mm at insertion) and more conspicuously spinose-toothed leaf margins. The upper portions of leaves in D. dicarpum are nearly filled by the costa and scarcely concave, whereas the upper portion of leaves in D. plurisetum are wider, more concave, and with laminae extending closer to the leaf apex. Although both species are plicate, the plicae of D. dicarpum are more distinct and usually extend further up the lamina. Under the microscope the distinctly shortened juxtacostal laminal cells in D. dicarpum entirely preclude confusion.
NI: N Auckland (Kaipara), S Auckland, Gisborne, Hawke’s Bay (Ikawetea Stream, Whirinaki River Track, Ruahine Range near Sunrise Hut), Taranaki, Wellington; SI: Nelson, Marlborough, Canterbury, Westland, Otago, Southland; St.
Australasian. Tasmania*, mainland Australia*, New Guinea*. Klazenga (2003, p. 93) also recorded this species from Taiwan and Vanuatu.
Growing mostly on litter, humus or gravel on the forest floor, but also on rotting logs and occasionally on tree trunks. Growing in a variety of native forest types, but particularly well developed and abundant in drier Fuscospora solandri s.l. forests, where it can be a major component of the ground cover. It is an abundant species in the drier (and often heavily deer-grazed) mid elevations of Canterbury L.D. Also occurring under exotic species including sycamore (Acer pseudoplatanus) and Pinus radiata. In wetter forests it shows a marked tendency to grow on well-drained sites and is absent from steep, damp banks. It is most abundant east of the Main Divide on the South I. but can be well developed in drier sites west of the Divide. Commonly associated with other Dicranoloma spp. (especially D. billardierei, D. fasciatum, and D. robustum) and Ptychomnion aciculare. On the North I. ranging from 50 m (Butterfly Creek, Wellington L.D.) to at least 1160 m (near Dawson Falls, Taranaki L.D.) and from low elevation (Coutts I., Canterbury L.D. and Pelorus Bridge, Marlborough L.D.) to at least 900 m (Magnesite Quarry on Cobb Reservoir Road, Nelson L.D).
There is a marked tendency in this species for stems to grow horizontally (as stolons) and give rise to erect secondary stems, often with elongate masses of aggregated rhizoids at branching points. Fertilisation apparently occurs in late summer, and in early winter "spear" stage sporophytes scarcely exceeding the perichaetial leaves can often be observed.
Material of this species from A.C.T., Australia (H. Streimann 5243, CHR 349542) has been seen in which the rhizoids give rise to masses of linear to fusiform, 6–9-celled, transversely septate gemmae. These rhizoidal gemmae are similar to those illustrated by Klazenga (1999) for several species of Malesian Dicranoloma. No rhizoidal gemmae have been seen in N.Z. material.
The presence of a large group of short juxtacostal cells is suggestive of the pan-tropical genus Leucoloma. However, in that genus the laminal cells are usually papillose and the costae have a very different structure from that observed in this species.
Dixon (1913, p. 20) treated "Dicranoloma leucolomoides" as a recognised species in the N.Z. flora, while Sainsbury (1955) included this name in synonymy of D. robustum. The Mossman collection from Kaipara Harbour selected as the lectotype of Dicranum leucolomoides Müll.Hal.by Klazenga (2003) is unquestionably referable to Dicranoloma dicarpum. However, duplicates of three of the other specimens cited by Dixon (1913) as D. leucolomoides are present in the Beckett herbarium at CHR and are referable to D. robustum. These specimens may have generated confusion surrounding the identity of Dicranum leucolomoides.
Dicranum dicarpum var. spinosum Hook.f. & Wilson was initially placed in the synonymy of Dicranoloma dicarpum by Dixon 1913, p. 14 and confirmed by Klazenga (2003). This synonymy is accepted here.
Brotherus (in herb.) applied the name Dicranum polysetum Hampe nom. illeg. to N.Z. material of Dicranoloma dicarpum. The Australian name Dicranum polychaetum Mitt. has not, to my knowledge, been applied to N.Z. material. These two names are not considered further here. Dixon (1913, p. 14) cited other Australian names (and their various combinations) in the synonymy of D. dicarpum; Klazenga (2003) verified these synonymies and typified them to the extent that he was able.