Class
Family
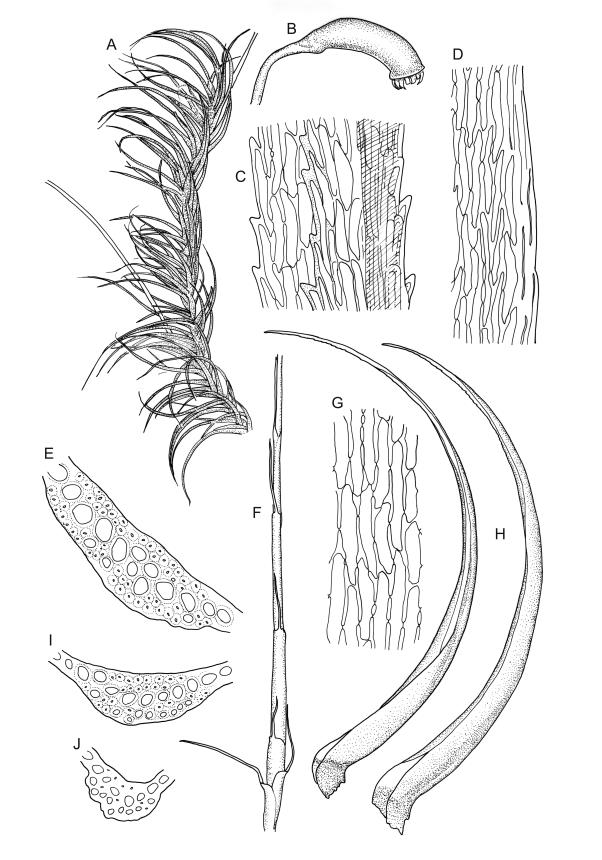
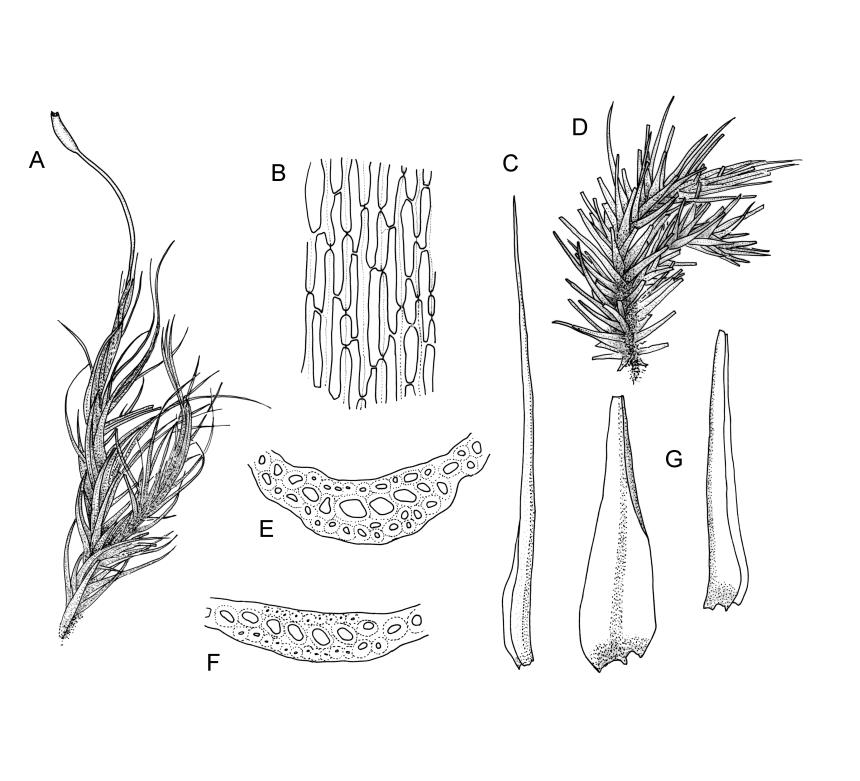
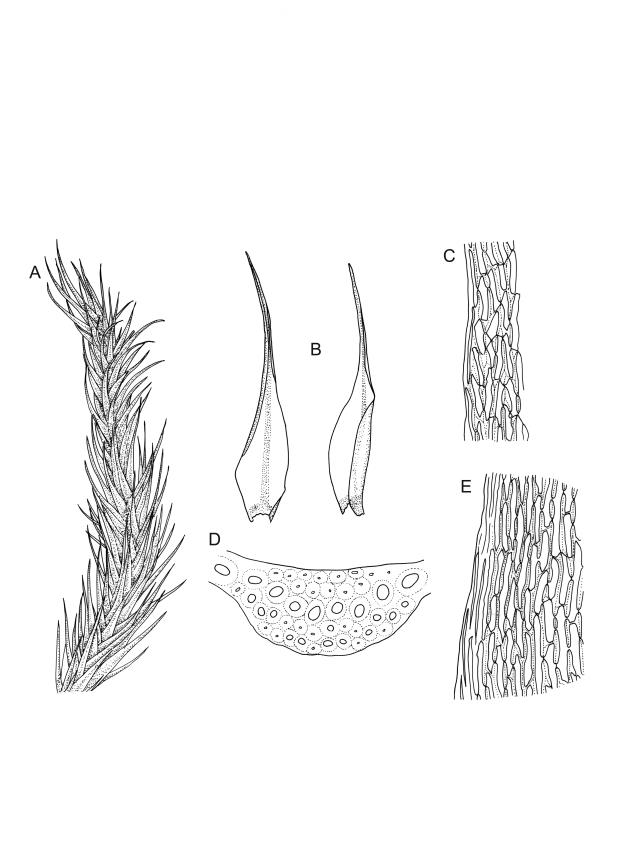
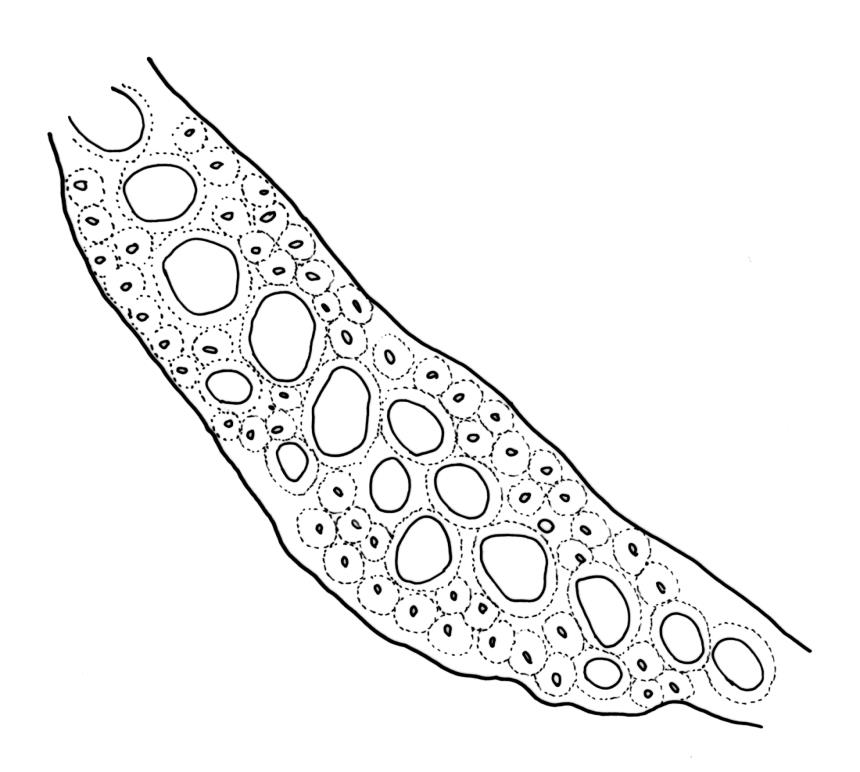
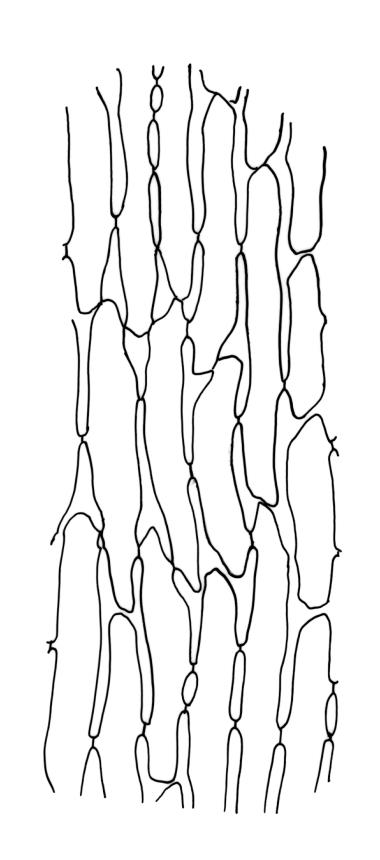
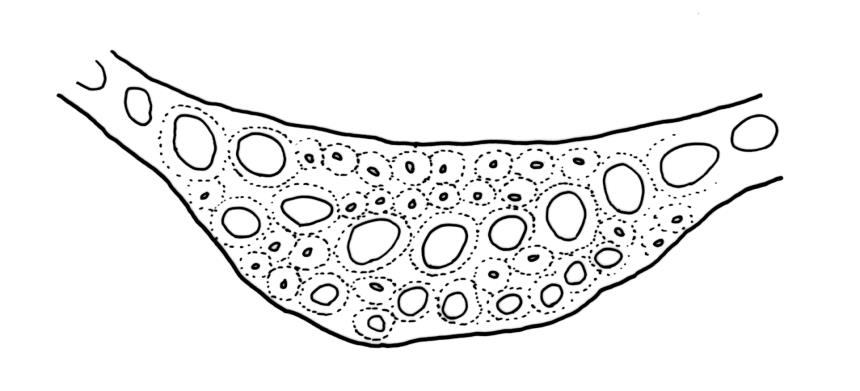
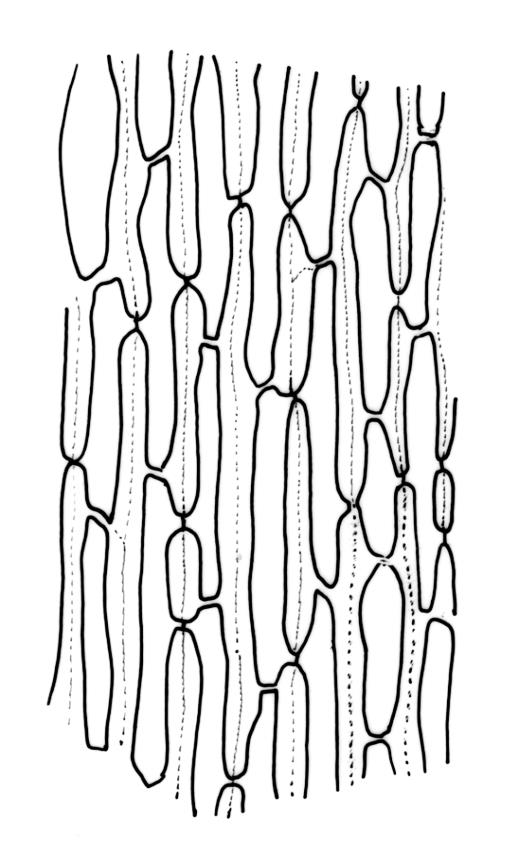
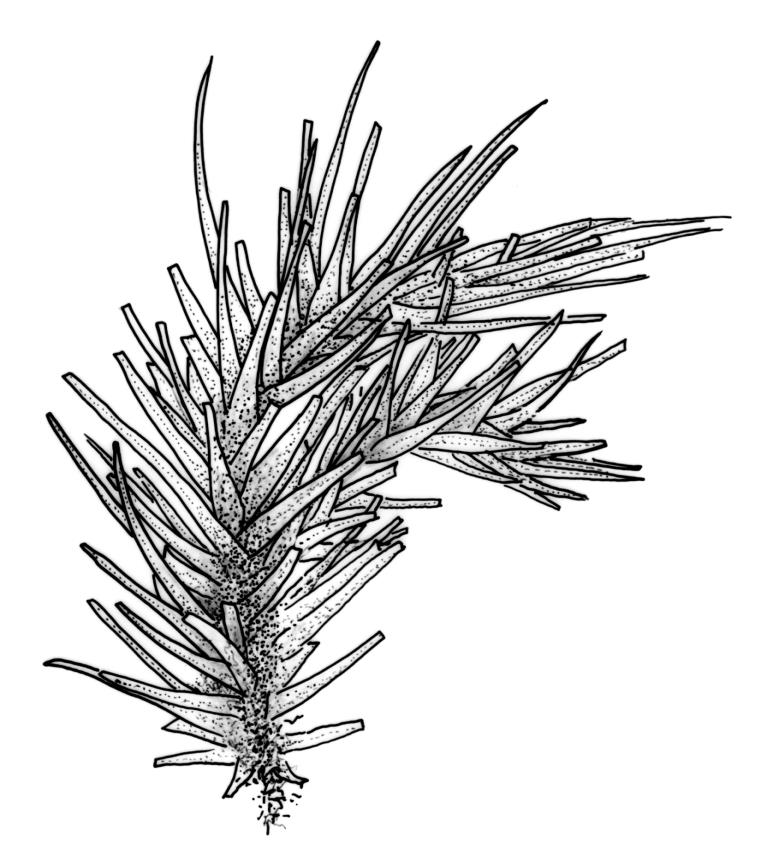
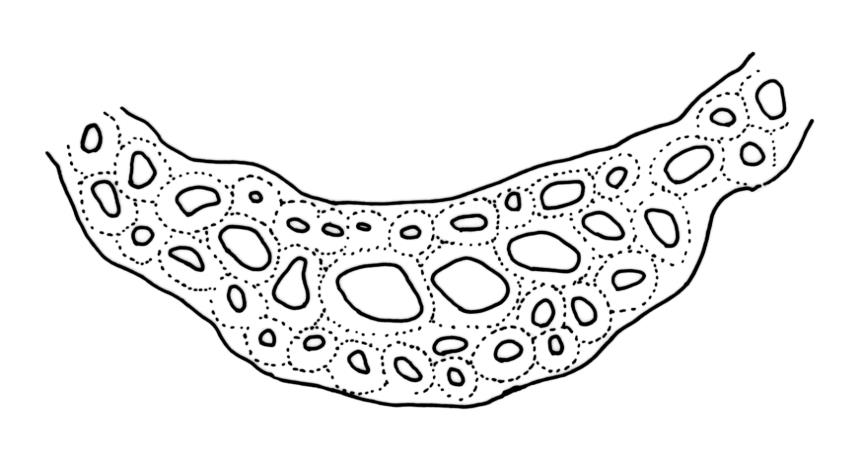
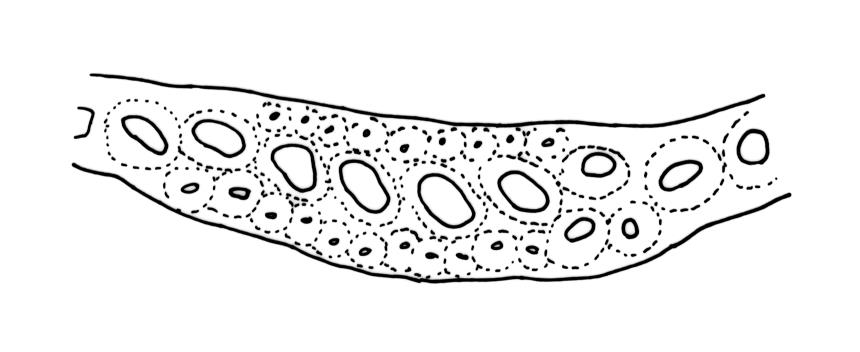
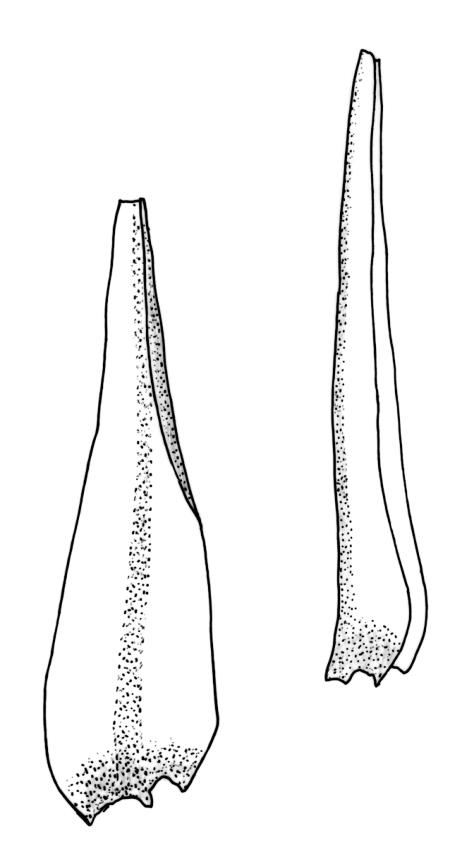
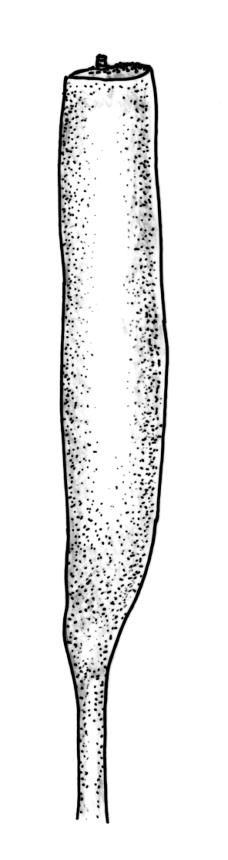
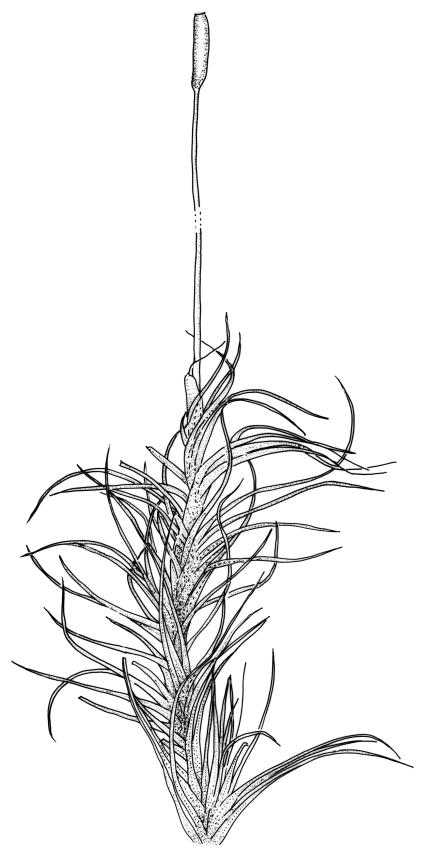
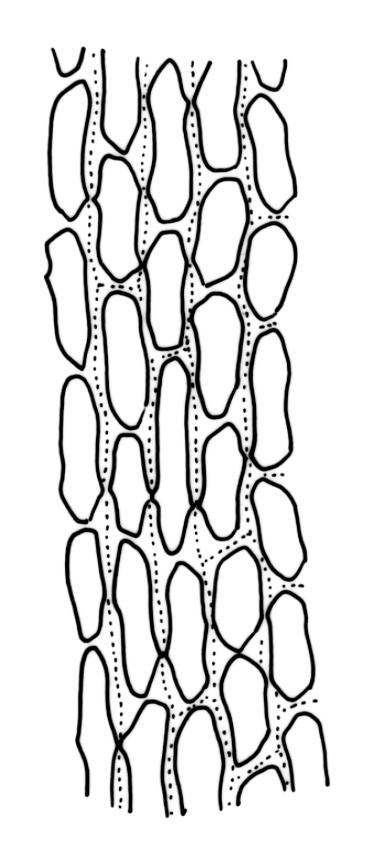
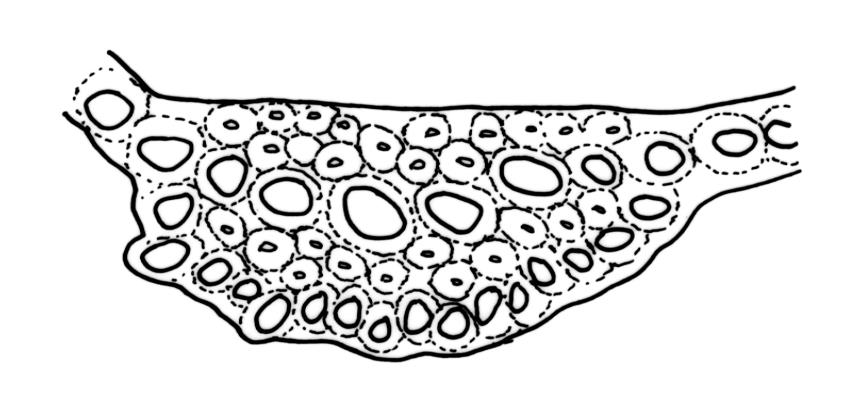
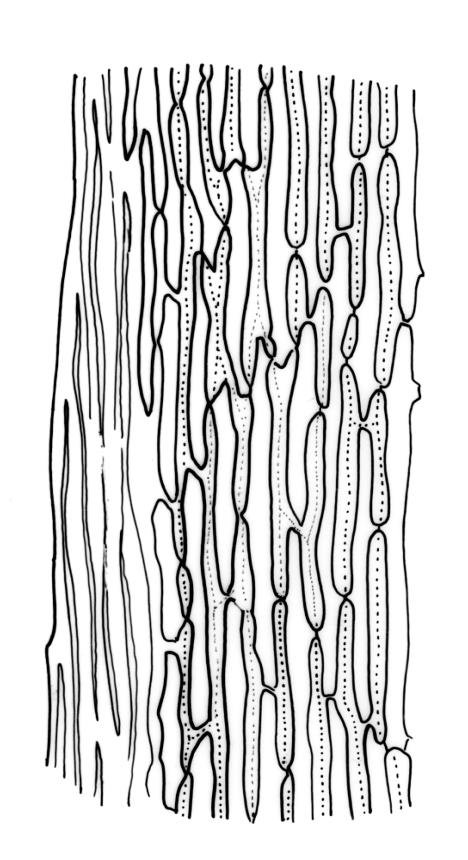
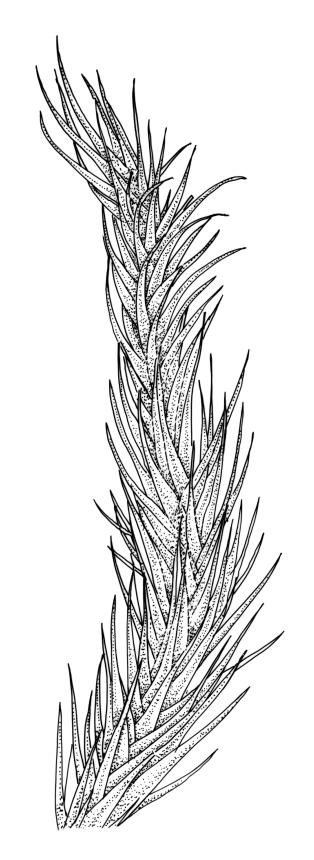
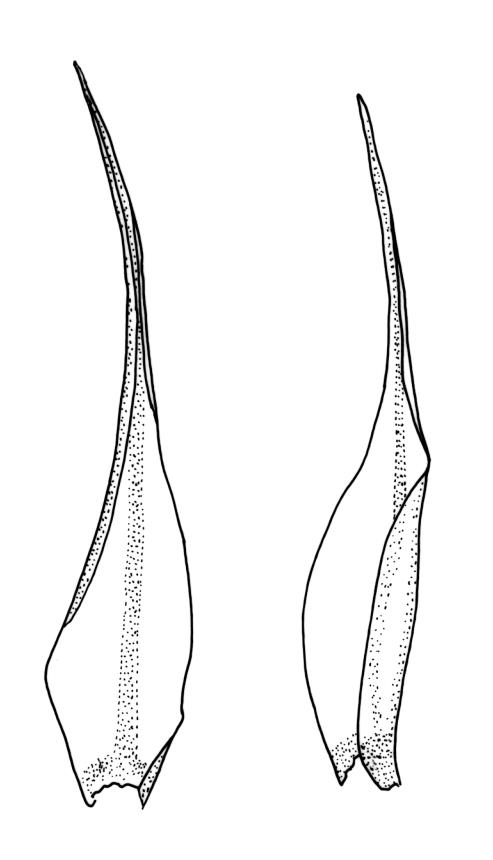
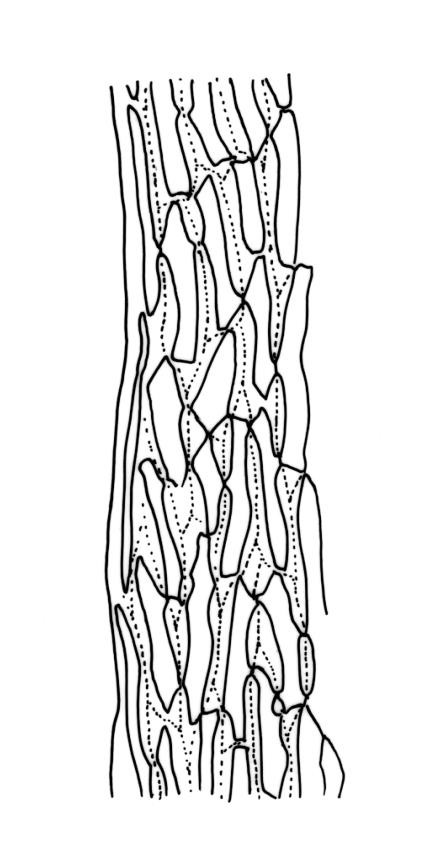
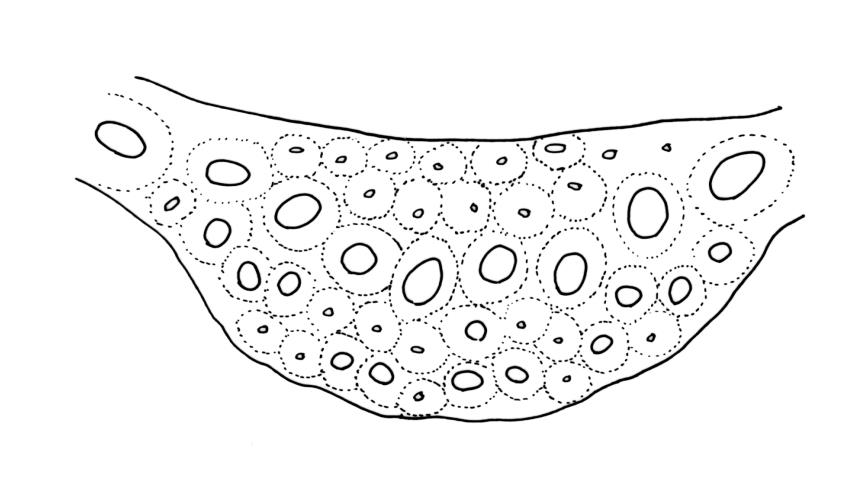
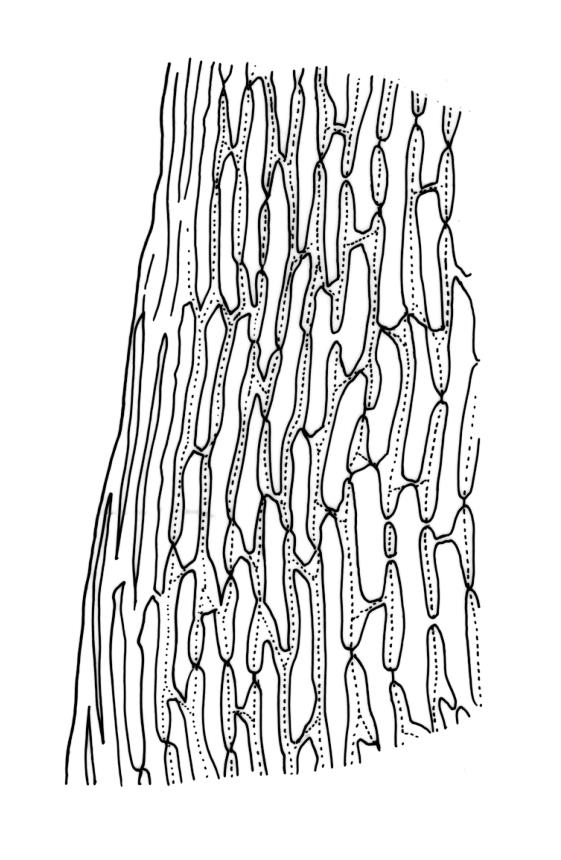
Plants robust, usually yellow-green and ± lustrous when fresh. Stems (20–)80–120 mm, sparsely branched by forking, in cross-section with 3–5 layers of cortical cells and a central strand; rather sparsely matted by pale rhizoids. Leaves falcate-secund or erect spreading, neither plicate nor rugose when fresh, little altered when dry, narrowly lanceolate, weakly toothed (not spinose) in upper ¼ to ⅓ or entire, (10–)12–14(–22) × c. 2.0 mm (under cover slip), clasping and ± auriculate at base, tubulose and plane at margins, often with tips fragmenting; mid laminal cells elongate and ± irregular, (45–)60–90(–12) × 9–12 µm and 5–14:1, thick-walled and highly porose, with elongate cells extending ± to apex; juxtacostal cells at mid leaf not differentiated; border mostly narrow (somewhat variable in width), extending from just above the alar group to lower limit of teeth (usually c. ¾ the leaf length), mostly 2–4 cells and 8–12(–15) µm wide at mid leaf; cell of leaf base longer, mostly 95–150 µm but otherwise as those of mid lamina, highly porose; alar cells strongly and abruptly differentiated, unistratose, ± auriculate, extending c. ⅔ to costal base and extending 15–20 cells up the margins, inflated and yellow-brown, firm-walled, the largest c. 70–90 × 45 µm. Costa narrow, occupying 1/20 or less of the widest part of leaf base, 90–105(–150) µm wide, weakly spinose abaxially above, in cross-section (at mid leaf) lacking wings, with a median layer (rarely 2 layers) of 6–8(–10) guide cells, a unistratose abaxial stereid group, a bistratose (often of 12–20 cells) adaxial stereid group (both extending the width of the costa), and a single layer of thinner-walled and often opaque cells (lumens c. 6 µm) on the abaxial surface.
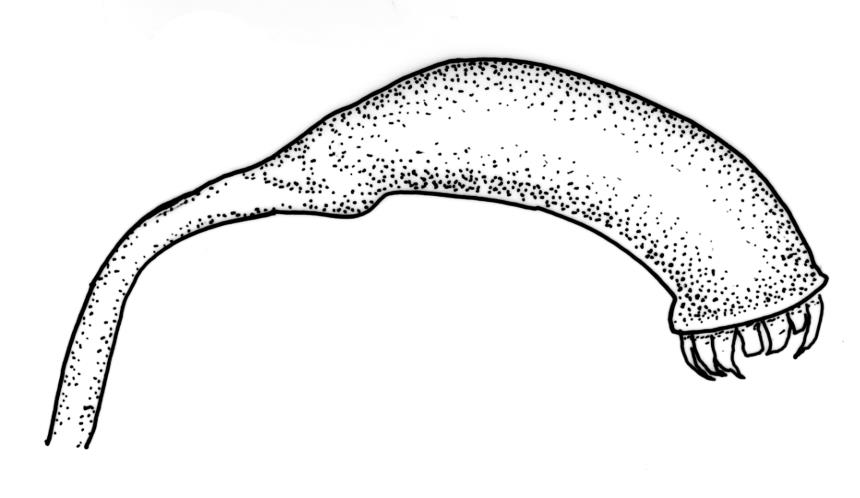
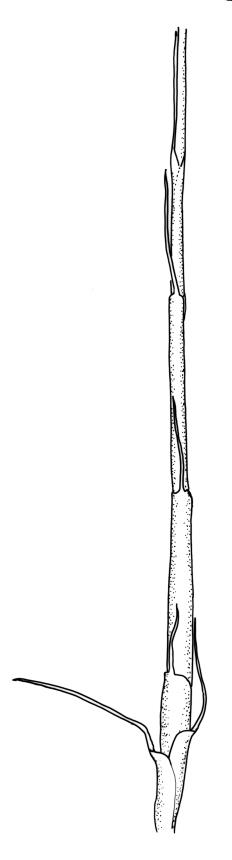
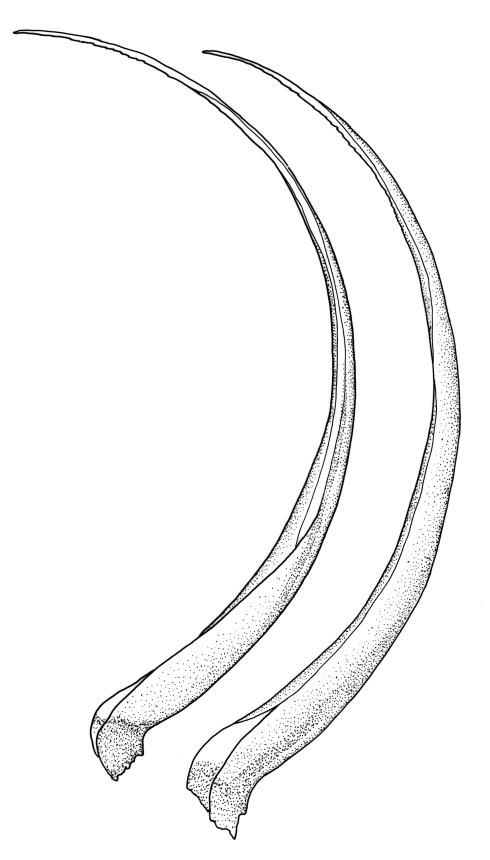
Pseudautoicous. Outer perichaetial leaves weakly costate or ecostate, with an oblong and strongly sheathing base abruptly tapered to an arista that is mostly equal to or longer than the base; inner perichaetial leaves ecostate, sheathing the bottom ⅓–½ (rarely more) of the seta, mostly 8–14 mm (including arista), gradually or abruptly tapered to a long arista (not formed by a costa) that is mostly 0.8–2.2 mm long. Dwarf males gemmiform, embedded in rhizoids of sterile or ♀ plants. Setae 1 per perichaetium, 14–27(–30) mm; straight, not twisted, pale brown; capsules exserted, curved, 2.3–3.5 mm, weakly to distinctly strumose, mostly smooth or nearly so and weakly constricted below the mouth when dry; exothecial cells incrassate, mostly oblong or elongate but some irregular, mostly 35–90 µm long; stomata restricted to neck, superficial; annulus weakly differentiated, not revoluble; operculum curved-rostrate from a conic base but often fragmenting, ± equal to the capsule. Peristome teeth inserted at mouth, red-brown, split ½–⅗ to base into segments of unequal width, c. 700 × 110 µm papillose-striolate in lower ½–⅔, baculate near apex. Calyptra cucullate, smooth. Spores spherical, 16–21 µm, nearly smooth.
Differences between Dicranoloma robustum and D. billardierei are discussed under the latter species.
NI: N Auckland, offshore islands only (LB), S Auckland, Gisborne, Hawke’s Bay, Taranaki, Wellington; SI: Nelson, Marlborough, Canterbury, Westland, Otago, Southland; St; Sol; A; Ant; C; M.
Austral. Tasmania*, mainland Australia*, Argentina*.
Mostly on duff and humus in southern beech forest and often forming extensive turves of many square metres. Also occurring on exposed roots, logs, tree trunks, and rock (and then usually as the "setosum" growth form), and occasionally on damp or waterlogged peat. In moist subalpine forest (such as at Arthur’s Pass), D. robustum is often the most conspicuous bryophyte and a dominant component of terrestrial bryophyte groundcover (but often mixed with various other species of Dicranoloma). Commonly extending into grasslands and into alpine habitats; D. robustum and the less common D. obesifolium are the only species of the genus commonly occurring in alpine grasslands. In lowland areas (below c. 400 m elevation) often associated with kānuka or mānuka forest scrub or on exotic trees such as poplars, eucalypts, or pines. The wide ecological tolerance of this species means that it associates with a wide range of other moss species, but most commonly, on forest floors, with other Dicranoloma spp. (especially D. billardierei, D. plurisetum, and D. dicarpum), Distichophyllum pulchellum, and Ptychomnion aciculare. In grasslands it very commonly occurs in association with the lichen Cladia aggregata.
Dicranoloma robustum is among the most widely distributed, common, and variable moss species in the N.Z. flora. Its apparent absence from N Auckland L.D., with the exception of occurrence on Little Barrier I., is curious. The description above applies to the most common forest floor growth form occurring on duff and humus through much of the country and conforms to the Hermite I. type. This representative growth form is a readily recognisable species by virtue of its robust habit, non-plicate and markedly secund leaves, solitary sporophytes, and long-aristate inner perichaetial leaves. In this most common growth form, the internal structure of the mid leaf costa is fairly constant and conforms to the above description, as well as to Renauld’s (1909) "toxoneuron" type. A single description attempting to encompass its full range of variability would require so many qualifications as to render it of little use.
Dicranoloma robustum is disturbingly variable and is necessarily interpreted broadly here. The broad concept employed here is unsatisfying and there is a lingering sense that the species is a taxonomic complex, which might be further resolved with a detailed study of patterns of morphological variation. However, both past and present efforts to do so have met with little success. A disturbingly large number of specimens/populations are morphologically intermediate between the most distinctive growth forms (discussed below) of this species. The large number of intermediates suggest they are environmentally induced growth forms, and they are here considered to be unworthy of taxonomic recognition.
Costal anatomy and leaf shape, size, degree of fragility, and toothing, all vary in apparent response to substrate and other environmental factors. Several variations, considered here to be growth forms, have been taxonomically described. The most distinctive of the growth forms, and the most commonly applied names, are described briefly below.
The name D. robustum var. setosum (Hook.f. & Wilson) Sainsbury, in its various combinations, has been widely applied to N.Z. populations. The material so-named in N.Z. herbaria are plants with erect-spreading, non-secund, narrowly setaceous, fragile leaves 7-15 mm long. Such material usually occurs on exposed roots, logs, tree trunks, and rock rather than on organic debris. Costal internal structure and width are quite variable in the "setosum" growth form, as is the degree of marginal toothing and border development. Populations in which the leaves are setaceous and fragile but markedly secund are common and widespread. Both Dixon (1913, p. 17) and Sainsbury (1955, p. 130) noted that in many populations of the "setosum" growth form, the number of layers of stereids surrounding the median guide cells (in mid leaf cross-section) is reduced to a single abaxial and single adaxial layer, and the costa projects only weakly on the abaxial surface, thus corresponding to the "heteroneuron" costal type of Renauld (1909). In the majority of N.Z. populations of the "setosum" growth form, the costa at mid leaf is 90-110 µm wide, with (6-)8 median guide cells, but populations with narrower costae and as few as 4-5 guide cells also occur. There are no clear morphological boundaries that distinguish the "setosum" growth form from more typical populations of D. robustum. What is treated here as the "setosum" growth form is found throughout the full N.Z. geographic range of D. robustum.
When well developed, the "setosum" growth form has a very distinctive facies and it has been accorded both species and varietal rank. This taxon was described (as Dicranum setosum Hook.f. & Wilson) using syntypes from Auckland and Campbell Is. gathered by members of the Erebus & Terror Expedition of 1839-1843. There are three (or doubtfully four) such collections in the Hooker herbarium. The protologue includes a description of the sporophytes and leaves ("foliis erecto-patentibus strictis longissimis lanceolato-setaceis fragilibus apice serrulatis nervo latiusculo subexcurrente"). The fruiting syntype that best conforms to the protologue (and which, fortuitously, most closely conforms to the concept applied by Dixon (1913) to N.Z. material) was collected by D. Lyall in the Auckland Is.; this material was designated as the lectotype by Klazenga (2003). The lectotype has fragile, narrowly lanceolate-setaceous and weakly toothed leaves, mostly 9-12 mm long with mid laminal cells mostly 39-105 µm long; a border 12-15 µm wide; and a costa at mid leaf c. 120 µm wide, with 8-10 median guide cells, and ± unistratose stereids.
Dicranum pungens Hook.f. & Wilson was correctly placed in the synonymy of D. robustum by Sainsbury (1948) and confirmed by Klazenga (2003). Sainsbury noted Dixon’s (1913, p. 21) earlier comments that he had "felt very doubtful about retaining this as a species". The type collection from the Auckland Is. consists of robust plants with stems to c. 80 mm and secund, non-fragile leaves mostly c. 10 mm. The costae are more weakly developed than in more representative D. robustum, being at mid leaf c. 75 µm wide, with 6 median guide cells and abaxial and adaxial stereid bands that are 1(-2) cell layers thick. The single layer of thinner-walled and often opaque cells on the abaxial surface that is often present in D. robustum is absent. Dixon (1913, p. 21) indicated the costa in D. pungens to be "heteroneuron" in structure. I concur with Sainsbury’s (1955, p. 21) observations that the costae in D. robustum can sometimes be narrower and have reduced stereid bands ("heteroneuron" as opposed to "toxoneuron" anatomy).
There are numerous collections in herb. Beckett (CHR) named as Dicranum subpungens Hampe [Linnaea 30: 629, 1860] by Brotherus. According to Klazenga (2003) this name has several Victorian syntypes, from which he selected a lectotype. Klazenga’s placement of this name in synonymy is not questioned here.
Dicranoloma cylindropyxis Dixon was accepted at the species level by Sainsbury (1955, p. 132) and Beever et al. (1992, p. 47), but rejected by Klazenga (2003). It is rejected here. When well developed, this growth form of D. robustum is distinctive by its red coloration, long (c. 4 mm), nearly erect, cylindric, and scarcely strumose capsules, short (mostly <30 µm and c. 2-5:1) and weakly porose upper laminal cells, stout costa (c. 0.1 the width of the lower leaf) and rather narrow (c. 1.0 mm) leaves. In the type of D. cylindropyxis the perichaetial leaves sheath c. ⅔ of the seta length. The "cylindropyxis" growth form of D. robustum appears to be best developed and most common at mid elevations in Nelson and Westland L.D. However, all the above cited features appear to vary independently and no clear boundary can be discerned between the "cylindropyxis" growth form and the representative growth form of D. robustum.
Dixon (1913) cited two syntypes for D. cylindropyxis Dixon from the Müller herbarium; this material is probably non-extant. Neither of the two syntypes (Helms s.n. and Beckett 166) are present in the Dixon herbarium, although several paratypes are present there. A portion of the original fruiting Helms Paparoa Range collection in NY labelled "Dicranum cylindropyxis C.Müll. n.sp.", apparently in Helms’s script, was designated the lectotype by Klazenga (2003). There is ample fruiting material of T.W.N. Beckett 166 (from the Kelly Range, Westland L.D.) in the Beckett herbarium.
Type material of Dicranoloma grossialare Dixon was not found by Klazenga (2003, p. 467). Dixon clearly cited T.F. Cheeseman 66 from the Mt Arthur Plateau (Nelson L.D.) in the Müller herbarium at Berlin as the type. The holotype was probably destroyed in the WW2 bombing of the Berlin herbarium, and an attempt to borrow type material from BM has failed. There are two fruiting Mt Arthur specimens in herb. Cheeseman (AK 12159, AK 12161) under the name D. grossialare, both originally named as Dicranum robustum and later revised by Sainsbury. One bears the poorly legible Cheeseman number 68 (or 168?). Neither of the AK specimens fully agrees with the collection details in Dixon’s protologue and there is no evidence that either Müller or Dixon examined either. It seems likely that Dixon misquoted the collection number in his protologue. The habit and associated species of these two specimens suggest that they are duplicates of a single collection. AK 12159 is unremarkable given the variation of D. robustum. In the absence of material precisely agreeing with the protologue it is considered as likely type material. The penicillate nature of the upper leaves, cited in both the protologue and by Sainsbury (1955a, p. 133), is only weakly developed in this specimen. The leaves are c. 15 mm long, tubulose, and finely subulate in a manner suggestive of the "setosum" growth form of D. robustum. The mid laminal cells are mostly 45-60(-90) × 6-8 µm, thick-walled, and porose. The costa at mid leaf is c. 90-100 µm wide, with seven guide cells, and with well-defined two-layered stereid bands and larger-lumened abaxial cells. The tubulose nature of the leaves obscures the costa in some leaves, and this may have influenced Dixon’s description of the costa as "basin versus tenuis, saepe indistincta". In cross-sectioned leaves, however, the lower costa is c. 100 µm wide. The supposed indistinct nature of the lower costa is emphasised as a diagnostic feature by Sainsbury (1955, p. 132). A collection from Mt Hauhungatahi (Wellington L.D., G.O.K. Sainsbury 489, CHR 545962), named by Dixon as D. grossialare, and cited by Sainsbury (1955, p. 133), is likewise referable to D. robustum. This material more clearly exhibits the faint lower costa cited by Dixon, but the shoots are not apically penicillate. The lower costa here is wide (c. 90 µm), indistinct, and "heteroneuron" in structure. The variability of costal structure in these specimens thus mirrors that in D. robustum, especially in the "setosum" growth form.
Dicranoloma integrifolium Dixon was initially synonymised by Sainsbury (1948) and confirmed by Klazenga (2003). This name has been applied to N.Z. collections of D. robustum in which the leaves are decidedly less secund, shorter (c. 5-8 mm), less strongly bordered, and more weakly toothed than representative material. Such populations are widespread and common. Type material of this name in BM was cited by Klazenga (2003) but has not been seen for this study; there appears to be none of the relevant R. Brown collection in any N.Z. herbarium. However, examination of non-type material named by Dixon (e.g., G.O.K. Sainsbury 590 from Mt Arthur, Nelson L.D., CHR 541110) corroborates Sainsbury’s (1948; 1955, p. 129) opinion that this name belongs in synonymy. In this collection the leaves are mostly erect-spreading but become weakly secund at the stem apex, c. 5-6 mm, and weakly serrate to almost entire above. The mid laminal cells are relatively short (mostly 30-45 µm), thick-walled, and porose; the border c. 12 µm wide; the costa is excurrent and in mid leaf cross-section c. 100-120 µm wide, with eight guide cells, and with the stereid bands usually associated with D. robustum. I concur with Sainsbury (1955, p. 129) that populations of D. robustum with leaves only weakly toothed are not uncommon throughout its N.Z. range, and that such material is best considered an environmentally induced growth form. This growth form appears to be related to high levels of insolation and to occur on thin soil over rock and on dry peaty soils. On these bases D. integrifolium Dixon is included as a synonym here.
The name Dicranum integerrimum Broth. & Geh. in Broth. [Öfvers. Finska Vetensk.-Soc. Förh. 37: 152, 1895], based on Tasmanian material from Sprent River, was applied to N.Z. material by Dixon (1913, p. 27). This name was typified and placed in synonymy of D. robustum by Klazenga (2003, p. 460).
Klazenga’s (2003) rejection of Sainsbury’s (1948) placement of Dicranum leucolomoides Müll.Hal. in the synonymy of Dicranoloma pungens is accepted here. Type material of Dicranum leucolomoides Müll.Hal. is referable to Dicranoloma dicarpum (Nees) Paris.