Class
Order
Family
Genus
Perennial herb to 0.05 m tall. Stems creeping and prostrate, sometimes ascending at tips, rooting at nodes; hairs eglandular, somewhat retrorse, uniform, also a few longer glandular hairs sometimes present. Leaf bud indistinct; leaves diverging while still small, opposite-decussate but sub-distichous on prostrate stems, spreading; lamina thin, broadly ovate to orbicular or reniform, 3–8 mm long, 5–12 mm wide, dull green above, pale green beneath; midrib and lateral veins visible; surfaces and margins with mixed long eglandular and glandular hairs; margin crenate or crenate-serrate; teeth in 2–5 pairs; apex obtuse to rounded; base truncate to sub-cordate; petiole 1–3 mm long. Inflorescence a terminal raceme, 50–300 mm long, sometimes resuming vegetative growth with opposite leaves after flowering; flowers distant, 5–20, all bisexual; minute, undeveloped flowers sometimes present in axils of lower bracts; bracts alternate, leaf-like; pedicels filiform, erect, often deflexed at fruiting, 15–40 mm long, with mixed short eglandular and longer glandular hairs all around. Calyx lobes 4, obtuse, 2.5–3.5 mm long, sub-equal, mixed glandular- and eglandular-hairy on outer surface and margins, glabrous within. Corolla 8–10 mm diameter; tube white and yellow, 0.5 mm long, < calyx, eglandular-hairy inside; lobes 4, blue, spreading, sub-equal, elliptic to orbicular, 3.0–4.5 mm long, rounded; nectar guides dark blue. Stamen filaments white, 2.5–3.0 mm long; anthers blue. Style glabrous, 2–4 mm long. Capsules and seeds not seen in New Zealand material.
The creeping growth form and solitary flowers on long filiform pedicels are distinctive. V. lilliputiana plants are similar in being prostrate and creeping and having blue flowers, but their leaves are very much smaller, greyish-green, and entire, and flowers are carried on very short stalks, each with a pair of bracts. V. jovellanoides plants are also prostrate and have similar leaves, but their flowers are in short racemes and the corolla is white. Plants of V. persica have a spreading growth form, blue flowers, and leaf-like bracts, but their leaves are generally slightly narrower than long, ovate, truncate, and toothed (rather than circular, sub-cordate, and crenate), their pedicels are stouter and shorter, and the anterior corolla lobe is usually distinctly paler than the other 3 lobes. In addition they set fruit, whereas V. filiformis plants are sterile in New Zealand. Plants of V. hederifolia have a similar growth form although not so strictly prostrate, and they differ in leaf shape and size, pedicel hairs in 1 row, distinctive cordate triangular fringed calyx lobes, smaller and paler corolla, and by producing capsules, which contain large, cup-shaped seeds.
When not in flower, V. filiformis might be confused with unrelated small creeping plants, especially Glechoma hederacea (Lamiaceae), which is locally naturalised throughout New Zealand. Glechoma stems are 4-angled and have longer hairs than V. filiformis stems. G. hederacea leaves are usually larger, more deeply cordate, and have deeper and rounder crenations. Glechoma flowers are sub-sessile in axillary clusters, and the corolla is long-tubed and 2-lipped.
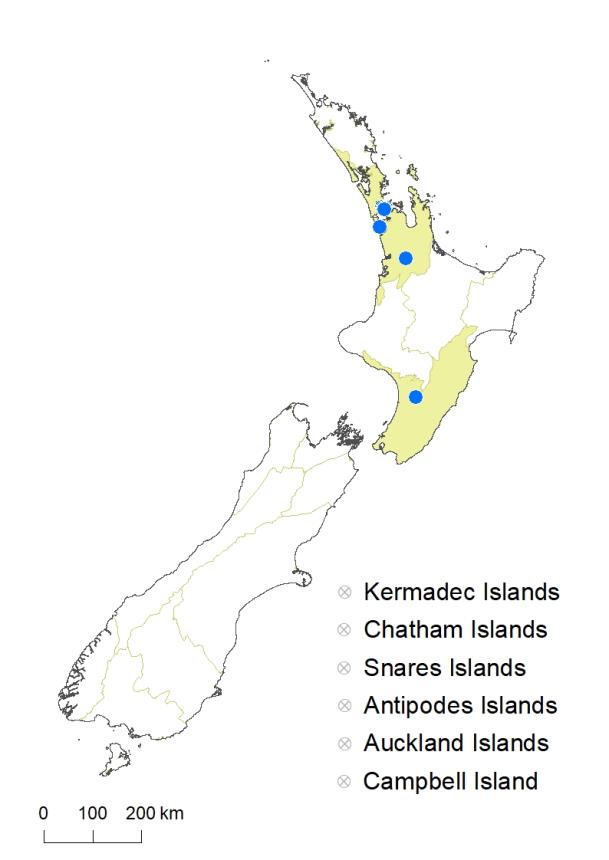
North Island: Auckland (in and near Auckland City, Hamilton), Southern North Island (Palmerston North).
Lawns, golf courses. Recorded elevations range from 30 to 120 m.
Indigenous to Caucasus, Crimea, and SW Asia.
Sykes (1981, p. 56). Voucher: CHR 363223, A.E. Esler, Mt Eden.
Flowers: August–November; fruits: not seen in New Zealand.
2n = 14 in Europe (Albach et al. 2008).
Veronica filiformis is classified in V. subg. Pocilla (Albach et al. 2004; Albach & Meudt 2010).
V. filiformis has spread around the temperate world as a cultivated plant and has escaped and become a troublesome weed in many places. The plants are self-incompatible and spread vegetatively, probably establishing freely from small fragments when lawns are mowed.
It is possible a single clone was introduced in New Zealand; in any case, self-incompatibility is the reason fruits are not produced in places where V. filiformis is naturalised. Scalone & Albach (2012) reported flower anomalies in clonal populations, which they attributed to accumulation of deleterious mutations in the absence of recombination and selection for flower function. The minute, undeveloped flowers seen in some New Zealand collections are likely to be of similar origin.