Class
Family
Genus
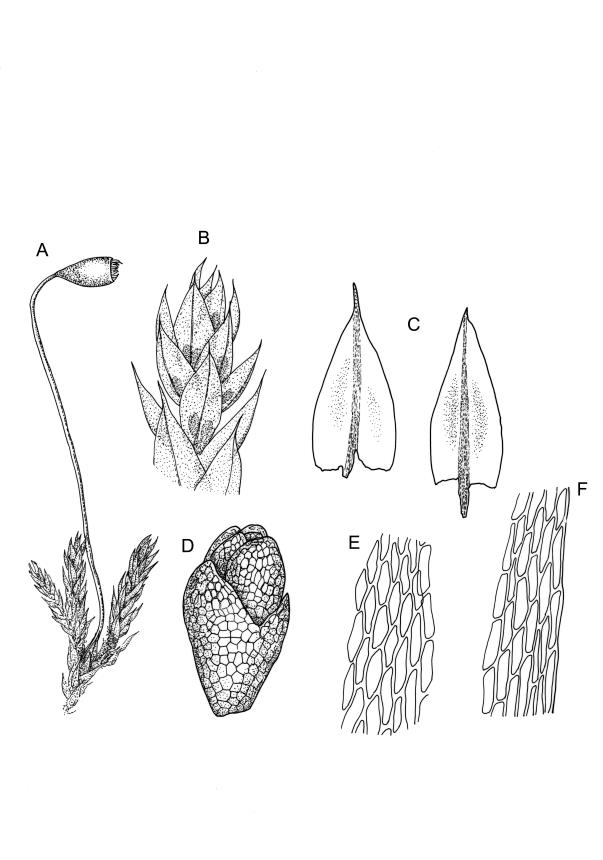
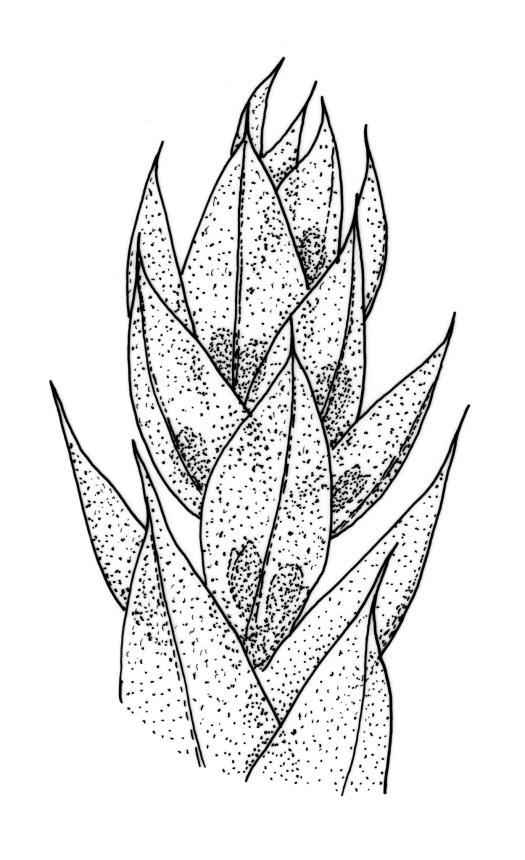
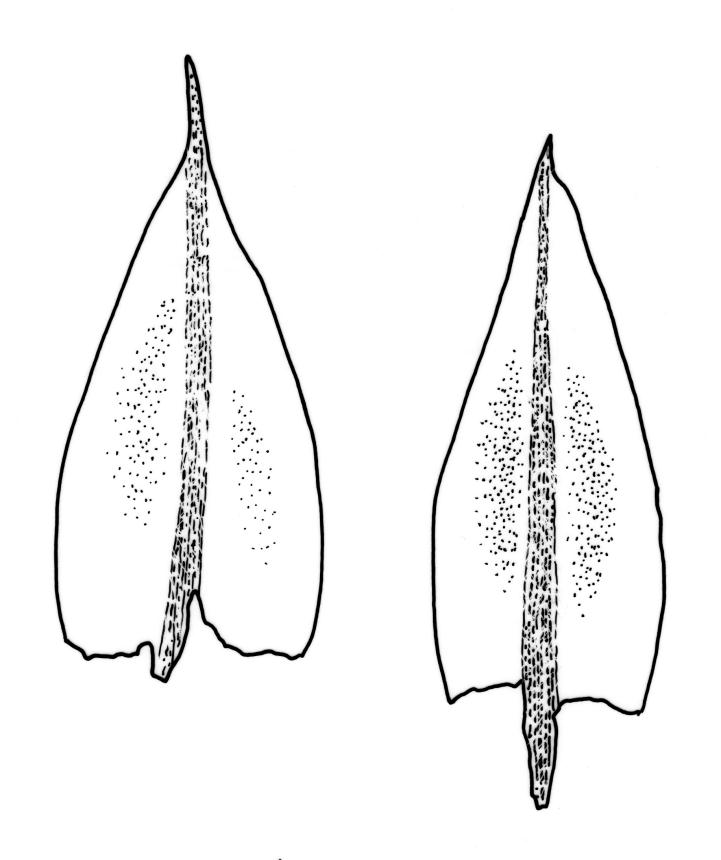
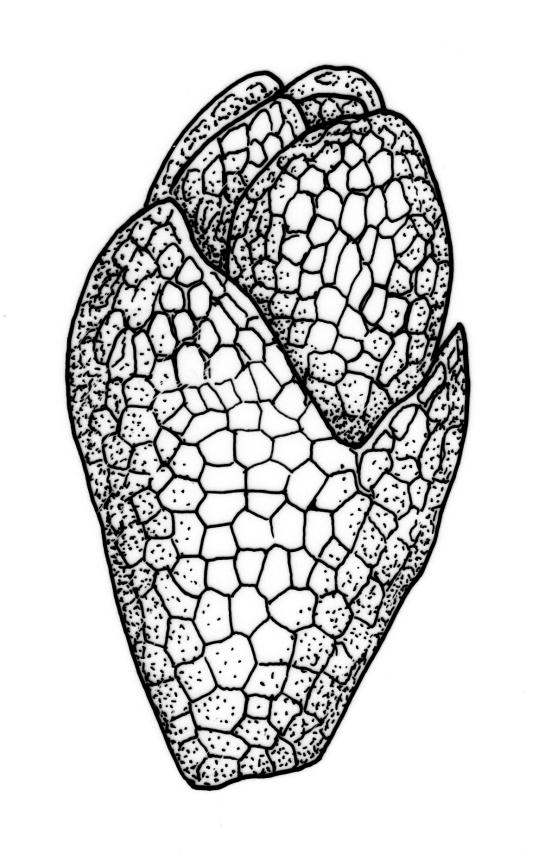
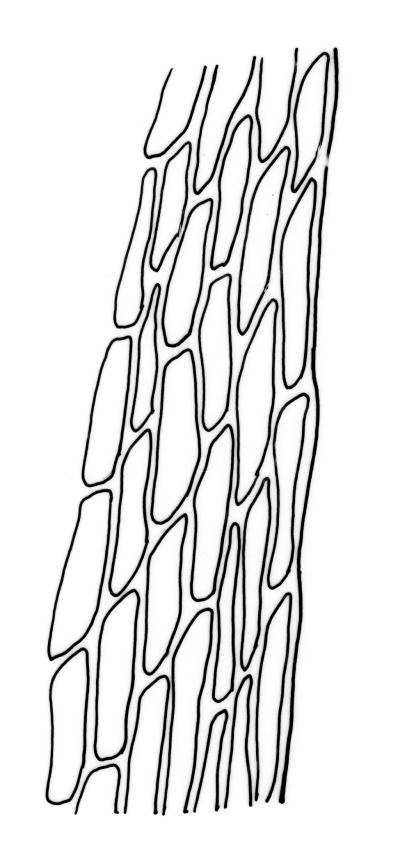
Plants yellow- or bright green, usually rather dull, forming loose turves on soil. Stems pale or red-brown, 7–15(–20) mm, sparsely branched, sparsely beset with red-brown, papillose rhizoids, in cross-section with firm-walled cortical cells and a distinct central strand. Leaves evenly spaced on stem, erect both moist and dry, ovate or ovate-lanceolate, acute, 0.7–1.4(–1.6) × 0.3–0.7 mm (lamina rarely >1.2 mm long) and with lamina 0.75–1.0 the total leaf length, concave, lacking secondary pigmentation, entire or nearly so, not or very weakly bordered and ± plane at margins, not decurrent; upper laminal cells hexagonal or rhombic, firm-walled, 27–39(–60) µm long and 3–5:1, becoming slightly shorter and more irregular near apex and more oblong in lower leaf; marginal cells somewhat longer (to c. 54 µm) and narrower in 2–4 rows to form a very weak and ill-defined border; basal cells short and oblong, not coloured. Costa stout, scarcely tapered, green or green-brown, short- or long-excurrent, or percurrent, usually forming a rigid, concolourous or apically hyaline, smooth or denticulate elongate cusp. Brood bodies (bulbils) usually present in upper leaf axils, obovoid or ellipsoid, variable in size (to c. 425 µm long) but mostly 1.5–2:1, with several primordial leaves occupying ⅓ to ½ the bulbil length; tubers absent (but see discussion below).
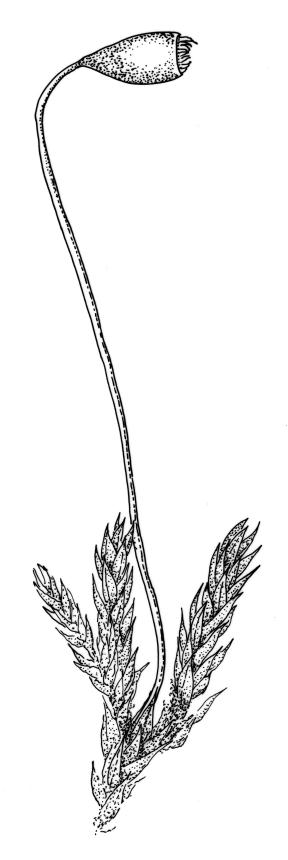
Dioicous. Perichaetia at base of plant; perichaetial leaves longer and more ovate-lanceolate and with costa more strongly excurrent (to c. ¼ the total leaf length). Perigonia terminal or overtopped by innovation, the inner bracts more concave and pigmented, but otherwise similar to vegetative leaves, enclosing antheridia and filiform (c. 6-celled) paraphyses. Setae c. 6–10 mm, red-brown, strongly curved just below capsule; capsules pendent or horizontal, obovoid, strongly constricted below the mouth when dry, c. 1–1.5(–2.0) × 0.7–1.1 mm, with an ill-defined concolourous neck c. ⅓ the total capsule length, as wide as or wider than the urn and strongly wrinkled when dry (poorly illustrated here); operculum lowly conic, not apiculate. Exostome teeth pale; endostome with narrowly perforate segments, ± equal the teeth, and moderately well developed, paired, and nodose cilia. Spores c. 15–18 µm.
Hedwig 1801, tab. 42, figs. 8–12; Ochi 1970, figs. 5–7; Seppelt 2004, fig. 38; Spence & Ramsay 2006, fig 42, x–ee (as Gemmabryum dichotomum); Ochyra et al. 2008, fig. 211.
When fruiting, the extremely short, obovoid, horizontal to pendent capsules provide distinction for B. dichotomum from other congeneric N.Z. species, with the exception of B. coronatum. My experience is that the capsule neck in B. dichotomum can be quite rough when dry, in contrast to the observations of Ochi (1970, p. 20) that it is "relatively smooth and never corrugate". This is not accurately illustrated in Image: Image\1ZL1, A. In B. dichotomum, however, the capsule neck in fresh material is concolourous with the urn at maturity, distinguishing it from B. coronatum, where the neck is usually much darker than the urn. Microscopically, the width of the endostomal segment perforations (narrow in B. dichotomum vs fenestrate in B. coronatum) and spore diameter (c. 15–18 µm in B. dichotomum vs 8–10 µm in B. coronatum) facilitate distinction between these two species. The nature of the leaf margins, the degree of secondary pigmentation of the leaves, and the presence of primordial leaves on bulbils also provide helpful distinctions from B. coronatum.
Confusion can occur (with sterile material) with B. clavatum, but the very weak leaf border and plane or nearly plane margins in B. dichotomum allow distinction in all but a few instances. When fruiting, capsule morphology readily distinguishes the two species.
Confusion sometimes occurs between B. dichotomum and B. sauteri; both form compact, bright yellow-green turves when sterile, and often occur in coastal situations. Like B. dichotomum, B. sauteri has stout, usually excurrent costae, and firm-walled upper laminal cells, but the more strongly bordered and recurved leaf margins, and the nearly constant presence of rhizoidal gemmae in B. sauteri allow separation in most instances. Also, B. sauteri lacks bulbils and has pyriform capsules with the neck narrower than the urn, which is a useful character when it is fruiting. In my experience, B. dichotomum lacks rhizoidal tubers. However, tubers (differing in morphology from those of B. sauteri) are recorded and illustrated from this species in Britain (Porley 2008, p. 61). J. Beever (pers. comm. 5 Mar. 2009) informs me that she has seen tuber-bearing material of B. dichotomum from N.Z. Spence & Ramsay (2006, p. 301) stated that "red globose [tubers are] sometimes present" in this species in Australia. Gall-like swellings on rhizoids also occur in some populations of B. dichotomum.
Bryum appressifolium, sometimes confused with B. dichotomum, is a more lustrous plant with leaves erect-imbricate and strongly recurved leaf margins.
K; NI: N Auckland, S Auckland (Te Kowhai, Coromandel Peninsula, Kaingaroa Plains), Gisborne (Arakihi, Marumaru, Lake Waikaremoana), Hawke's Bay (Ruakituri Valley, Napier), Taranaki (Mt Messenger), Wellington; SI: Nelson, Marlborough (Waima River), Canterbury, Westland (Franz Josef Glacier, Fox Glacier), Otago, Southland (Doubtful Sound); Ch. The above distribution is based on fertile collections only and undoubtedly understates the occurrence of this species.
Cosmopolitan. Tasmania*, mainland Australia*. Ochi’s (1985) assessment that this species is "widespread in every continent throughout the world" is accepted here. The range given by Ochyra et al. (2008, p. 464–467) for the "whole B. dichotomum complex" accords with Ochi.
On bare soil generally, including road cuts, sand dunes, and coastal banks; especially common on calcareous substrates. Also occurring in urban situations (footpaths, disturbed soil, etc.). On South I. this species ranges from sea level to at least 730 m. Frequently associated moss species include B. laevigatum, Ceratodon purpureus, Desmatodon convolutus, Funaria hygrometrica, Philonotis tenuis, Tortula muralis, and Weissia controversa.
Bryum dichotomum is a member of the so-called "B. bicolor complex" (sensu Wilczek & Demaret 1976).
The name B. dichotomum has been used as a catch-all for difficult Bryum material by many N.Z. botanists, and collections assignable to at least (but probably more than) three species have been incorrectly assigned to this species. Even with the removal of the clearly misnamed material, the morphological variability of this species is perplexing and other allied species (of the "B. bicolor complex") may be present.
Sterile material that is probably referable to B. dichotomum (e.g., B.H. Macmillan 89/140 ex Christchurch, CHR 413677) tends to bright yellow-green, with stems usually c. 10–14 mm and plants often rather lax in appearance. The leaves have plane, unbordered margins; green and percurrent to excurrent costa (usually more excurrent near the stem apices); green leaf bases; and abundant bulbils. While such material has a characteristic facies, a large proportion of sterile material cannot be named confidently and sterile collections are not incorporated into the above distribution. Several sterile collections from Marlborough L.D. can be only tentatively named. Sterile material from St, Sol, A, and C, differs from the usual form by having distinctly but narrowly recurved, bordered leaf margins.
A sterile collection from the Longwood Range, Southland L.D. (W. Martin, 1 Sep. 1950, CHR 515105), allied to the B. bicolor complex, is of special interest. The presence of distinct leaf borders, decurrent leaf bases, and unusual bulbil morphology precludes its placement in either B. dichotomum or B. coronatum. The material has stems 10–15 mm, with leaves that are ovate-lanceolate to lanceolate, c. 1.2–2 mm long, plane at margins, distinctly bordered (c. 4 cells wide at mid leaf), and decidedly decurrent. The upper laminal cells are firm-walled and mostly 30–45 µm long. The upper leaf axils bear 3–5 broadly obovoid bulbils, each c. 235–290(–380) × 140–150 µm, and with 3–4 leaf primordia occupying c. ½ their total length. This material may represent a species previously unrecorded from N.Z. or be undescribed, but it is too sparse to justify formal description.