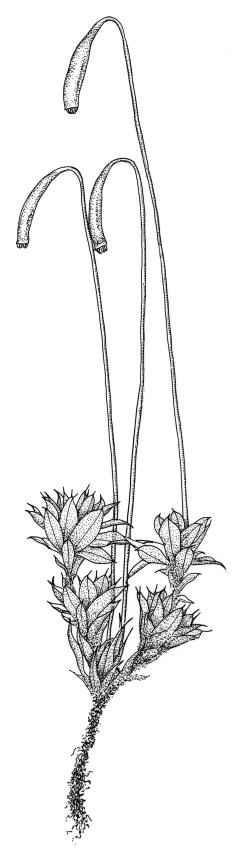
Class
Classification
Subordinate Taxa
Nomenclature
Scientific Name:
Bryaceae Rchb., Bot. Damen. 260 (1829) – as Bryoidea
Type Taxon:
Description
Plants small to robust, erect, extremely variable in size and coloration, mostly forming turves or tufts, perennial, terrestrial or occasionally epiphytic. Stems erect, from a few millimetres to several centimetres, usually strongly beset with rhizoids, in cross-section with firm-walled cortical cells and a distinct or rarely weak central strand of narrower thin-walled cells. Leaves evenly spaced or becoming larger and crowded above (comose), not complanate, spreading or erect-spreading when moist, often erect when dry, ovate, obovate, spathulate, elliptic, lanceolate, or ± cochleariform, acute, acuminate, or rarely rounded at apex, ± concave, often with secondary pigments, margins toothed or entire, mostly bordered and recurved, occasionally decurrent; upper laminal cells hexagonal or rhombic-hexagonal, thin- or firm-walled, not thickened at corners, porose or not, becoming more oblong in lower leaf; marginal cells mostly prosenchymatous and forming a distinct border of variable width and height. Costa strong, mostly excurrent but occasionally percurrent or shorter, with a single abaxial stereid band. Brood bodies (gemmae, tubers, or bulbils) often present.
Dioicous or monoicous. Perichaetia terminal or on short basal shoots. Perigonia mostly similar to perichaetia, with numerous filiform paraphyses. Setae elongate, single or occasionally multiple, often strongly curved just below the capsule; capsules horizontal, cernuous to pendent, rarely ± erect, pyriform, clavate, cylindric, or rarely ± globose, straight or curved, mostly with a well-developed neck; stomata numerous, superficial and restricted to neck; annulus well developed and revoluble; operculum mostly conic. Peristome double, often perfect; exostome teeth yellow-brown or pale, finely papillose below and mostly hyaline and baculate above, bordered, densely trabeculate on adaxial surface; endostome arising from a high or short basal membrane, with well-developed and perforate segments, and with cilia in groups of 1–3, these mostly appendiculate or nodose, rarely reduced or absent. Calyptra cucullate and smooth. Spores spherical and usually <30 µm.
Taxonomy
The Bryaceae are a large, cosmopolitan, and taxonomically difficult family. Brotherus (1924, p. 375) estimated that about 800 species belonged in Bryum alone, with it being by far the largest genus. The numbers of accepted species have reduced considerably and the accepted concept of the Bryaceae has changed greatly in recent decades. The Bryaceae are considered here to include (regionally) three genera: Bryum, Plagiobryum, and Rosulabryum.
Crum & Anderson (1981) in their treatment for eastern North America presented a broader and more traditional concept of the family than the one presented here. Their concept of the family included the genera Pohlia, Leptobryum, Mielichhoferia, and Orthodontium and largely followed the classification of Brotherus (1924). The treatment of the family by Smith (1978), in the first edition of his Moss Flora of Britain and Ireland, also largely followed Brotherus (1924) in his definition of the family (and its subfamilies). In the second edition of his Flora (Smith 2004) he presented a narrower and substantially different concept of the family.
The Bryaceae are conceived here in a fairly narrow sense, largely in accordance with Goffinet et al. (2009). Recent changes in the definition of this family and the generic limits within it are due mostly to the incorporation of molecular analyses (e.g., Pedersen et al. 2007; Cox & Hedderson 2003); such analyses, and their consequent changes in concepts, will undoubtedly continue in the future.
A recent treatment of the Bryaceae for Australia (Spence & Ramsay 2006) recognised eight genera in this family, of which three (Rosulabryum, Gemmabryum, and Ochiobryum) were recently described by one or both of the authors. Rosulabryum is accepted here, in the Bryaceae. Gemmabryum is not recognised in this Flora (see Excluded genera). Ochiobryum is accepted in this Flora but treated in the Mniaceae (see Excluded genera). The previously described Ptychostomum Hornsch. was resurrected by Spence (2005); it is not accepted here.
Rosulabryum was erected by Spence (1996), and Spence & Ramsay (1999) provided a key to the Australasian species. Rosulabryum was subsequently characterised by Spence & Ramsay (2006, p. 331) as comprising "the rosulate species of Bryum s. lat. with unreduced peristomes, generally obovate leaves with serrate margins, rhizoidal tubers and, occasionally, filiform gemmae in the leaf axils". They treated 14 spp. in the Australian flora in Rosulabryum, while four species are accepted here for N.Z.
For those wishing to apply Spence & Ramsey’s generic and species concepts, their keys, descriptions, and illustrations in the Flora of Australia are recommended.
Because of the complexity of the family, and differing opinions concerning familial, generic and species boundaries, all the species are distinguished here in a single key, given below.
Pohlia and Schizymenium (the latter roughly equivalent to Mielichhoferia sensu Brotherus) as well as Ochiobryum are here treated in the Mniaceae and Leptobryum in the Meesiaceae. These genera were all treated in the Bryaceae by Sainsbury (1955a), and many students of N.Z. mosses will attempt to name material by assigning them first of all to this family.
A practical rule-of-thumb, quoted here from Crum & Anderson’s (1981, p. 503) generic key, is that Bryum and its close allies have "leaves relatively broad, elliptic to ovate or ovate-lanceolate, usually distinctly bordered; cells rather broad, generally less than 4:1" whereas Pohlia, Leptobryum, and Schizymenium have "leaves linear to oblong-lanceolate, not distinctly bordered; cells narrow, relatively long, 4:1 or longer". This rule-of-thumb, written with American taxa in mind, is extremely useful but cannot be consistently applied to N.Z. taxa of Rosulabryum (which have ± spathulate leaves), Plagiobryum (which have rounded-hexagonal laminal cells and strongly bordered leaves), Ochiobryum (which have oblong-ovate and weakly bordered leaves) or Epipterygium (which has broadly elliptic and weakly bordered leaves).
Key
Key to the genera
| 1 | Upper laminal cells very thin-walled, not porose, and often lacking chlorophyll; leaves ± cochleariform, not bordered, broadly rounded at apex, evenly spaced and imbricate, c. 0.5–0.7 mm (excluding perichaetial leaves); costa ending far below leaf apex | Plagiobryum |
| 1' | Upper laminal cells mostly firm- or thick-walled, often ± porose, nearly always chlorophyllose (except in B. argenteum); leaves mostly ± ovate, ovate-lanceolate to spathulate or obovate, very rarely ± cochleariform (then larger and with firm-walled cells), often bordered, mostly acute or acuminate; costa mostly excurrent, less often percurrent or shorter | 2 |
| 2 | Leaves spathulate or obovate, widest at or slightly above mid leaf, nearly always comose when bearing sex organs (sometimes evenly spaced on sterile stems); mostly spirally twisted round stem or occasionally (in R. subtomentosum) spirally twisted around the leaf axis or little altered when dry | Rosulabryum |
| 2' | Leaves not widest above mid leaf, ovate, or ovate- to oblong-lanceolate, rarely oblong-spathulate to elliptic (in Bryum laevigatum and then not comose), or cochleariform, not or sometimes ± comose when bearing sex organs; erect or variably twisted when dry | Bryum |
Key to the species
In the following key plant size definitions follow Ochi (1970, p. 14) and are based on the length of the largest stem leaves (including perichaetial leaves), but excluding the excurrent portion of the costae. Robust: leaves >4.0 mm; large: leaves 3.0–4.0 mm; medium-sized: 1.5–3.0 mm; small: <1.5 mm.
| 1 | Bulbils (reduced bulb-shaped branches) ellipsoid or obovate, often present in axils of upper vegetative leaves; capsules <2 mm, when mature with necks as wide as or wider than the urn and strongly wrinkled when dry | 2 |
| 1' | Bulbils in axils of upper vegetative leaves very rare; capsules mostly >2 mm (excluding B. argenteum), when mature with necks narrower than the urn, moderately wrinkled or smooth when dry (excluding B. argenteum) | 3 |
| 2 | Leaves mostly with distinct red pigmentation; bulbils lacking apical primordial leaves; endostome segments fenestrate; spores 8–10 μm; operculum shortly apiculate | Bryum coronatum |
| 2' | Leaves lacking red pigmentation; bulbils with several apical primordial leaves; endostome segments narrowly perforate; spores c. 15–18 μm; operculum not apiculate | Bryum dichotomum |
| 3 | Plants hoary when dry; costa failing far below the leaf apex; upper laminal cells colourless, usually markedly thinner-walled than cells of lower leaf, rarely not differentiated (and then leaves broadly rounded apically) | 4 |
| 3' | Plants not hoary when dry; costa nearly always percurrent to long-excurrent (except in B. harriottii); upper laminal cells with chlorophyll, not markedly thinner-walled than cells of lower leaf | 5 |
| 4 | Leaves mostly acute, shortly apiculate or acuminate, only occasionally obtuse to rounded at apex; laminal cells near leaf apex and upper margins markedly thinner-walled than in lower leaf; capsules 0.85–2.0 mm, pendent, obovoid to oblong-cylindric, with poorly defined necks c. ¼ the total length and wrinkled when dry; spores 9–15 µm, smooth, lacking trilete scars; plants very common and weedy in wide variety of terrestrial habitats | Bryum argenteum |
| 4' | Leaves consistently cochleariform and broadly rounded at apex; laminal cells near leaf apex not clearly differentiated from cells in lower leaf, thin-walled throughout the leaf; capsules elongate and curved, c. 5–6 mm, horizontal or nearly so (sometimes pendulous when dry), with narrow necks c. ½ the capsule length and not markedly wrinkled when dry; spores 27–33 µm, papillose, often with trilete scars; plants less common, not weedy and usually associated with base-rich rock | Plagiobryum novae-seelandiae |
| 5 | Leaf apex broadly obtuse or rounded (Ochiobryum blandum will also key here; it has upper laminal cells thin-walled and mostly 5–7:1; delicate costae, cerise or black colouration (at least in part), and is aquatic or nearly so; it is treated here in the Mniaceae) | 6 |
| 5' | Leaf apex acute or acuminate | 7 |
| 6 | Plants julaceous, small; stems nearly always <10 mm; vegetative leaves c. 1 mm or less, broadly ovate to ± cochleariform; spores >20 µm | Bryum harriottii |
| 6' | Plants not julaceous, robust; stems often >20 mm; vegetative leaves >1.5 mm, oblong-spathulate to broadly elliptic; spores <20 µm | Bryum laevigatum |
| 7 | Plants robust and strongly comose, at least some usually plurisetose; well-developed leaves mostly >4 mm excluding excurrency, with apices mostly strongly reflexed; setae with a characteristic hook below the capsule | Rosulabryum subtomentosum |
| 7' | Plants not robust, comose or not, not plurisetose; well-developed leaves <3.2 mm excluding excurrency, with apices not strongly reflexed (leaves rarely to 5 mm, but then with apices not reflexed); setae not hooked below the capsule | 8 |
| 8 | Leaf border in widest part of leaf ≥10 cells wide; leaves comose, apically reflexed, spiralled around stem when dry | Rosulabryum perlimbatum |
| 8' | Leaf border in widest part of leaf ≤8 cells wide; leaves if comose not spiralled around stem when dry | 9 |
| 9 | Endostomal cilia well-developed and appendiculate; spores usually <20 µm . | 10 |
| 9' | Endostomal cilia not appendiculate (cilia may be absent or present and nodose or rudimentary); spores more variable in size, usually >20 µm (Rosulabryum campylothecium is likely to cause difficulty here. It is characterised by its golden colouration, serially comose habit, and strongly awned leaves little altered when dry. It mostly lacks capsules and is coastal. Its capsules are banana-shaped, with nodose cilia and spores ≤ c. 20 µm diam. Bryum preissianum may also cause difficulty here; it has no cilia or rudimentary ones but spores are 8–12 µm diam.) | 30 |
| 10 | Leaves closely imbricate; plants catkin-like, ≤7 mm high, very pale green | Bryum funkii |
| 10' | Leaves not imbricate (except in Bryum appressifolium); plants not catkin-like, usually larger, coloration various | 11 |
| 11 | Costa of upper leaves strongly excurrent | 12 |
| 11' | Costa of upper leaves percurrent, subpercurrent, or short-excurrent to form a cusp | 27 |
| 12 | Leaves decurrent at base (especially obvious in sterile plants); synoicous | Bryum pseudotriquetrum |
| 12' | Leaves not decurrent; mostly dioicous | 13 |
| 13 | Stems obscured by leaves; leaves densely erect-imbricate, with a single plication on one or both sides of the costa (best visible when leaves are removed from the stem); margins strongly recurved | Bryum appressifolium |
| 13' | Stems not obscured by leaves; leaves not densely erect-imbricate, lacking plications; margins variably recurved or plane | 14 |
| 14 | Leaves moderately to strongly spiralled around stem when dry; upper laminal cells thin- or firm-walled | 15 |
| 14' | Leaves not spiralled around stem when dry; upper laminal cells firm-walled | 17 |
| 15 | Leaves ± spathulate (widest above the middle and narrowed at base); upper laminal cells thin-walled; cilia markedly appendiculate; spores <15 µm; tubers often present | Rosulabryum capillare |
| 15' | Leaves ± ovate (widest below or near middle and not narrowed at base); upper laminal cells firm-walled; cilia appendiculate or sometimes merely nodose; at least some spores >15 µm; tubers absent | 16 |
| 16 | Synoicous; capsules ≤2.5 mm, narrowly pyriform; spores >20 µm; cilia paired and nodose to weakly appendiculate | Bryum pallescens |
| 16' | Autoicous or polygamous; capsules 2.5–3 mm, oblong-cylindric; spores mostly <20 µm; cilia mostly in 3’s, appendiculate | Bryum creberrimum |
| 17 | Leaves with a weakly to moderately reflexed cusp, not or sometimes weakly comose; tubers lacking | Rosulabryum billardierei |
| 17' | Leaves lacking a reflexed cusp, not comose; tubers often present | 18 |
| 18 | Excurrent part of costa mostly entire; capsules inclined to horizontal and distinctly curved in neck; spores >20 µm diam. | Bryum clavatum |
| 18' | Excurrent part of costa denticulate (rarely ± entire); capsules pendent, not curved in neck; spores <20 µm diam. (rarely c. 20 µm in Bryum sauteri) | 19 |
| 19 | Tubers present on rhizoids | 20 |
| 19' | Tubers absent from rhizoids | 26 |
| 20 | Tubers mostly 2 cells wide, pyriform or ellipsoid, mostly <100 µm in length, with cell walls not protruding; cilia mostly single ["B. erythrocarpum complex" pro parte] | Bryum sauteri |
| 20' | Tubers more than 2 cells wide, globose, oblong, or irregular, mostly >100 µm in greater diam. (or, if less, then irregular in outline and plants with entire leaf margins), with cell walls protruding or not; cilia paired or in 3s | 21 |
| 21 | Tubers irregular in outline, (60–)70–90(–105) µm in greater diameter; leaf margins entire; spores mostly >20 µm | Bryum clavatum |
| 21' | Tubers mostly globose, occasionally oblong, or ± irregular in outline; leaf margins mostly denticulate (costal excurrency, at least, always denticulate); spores 9–15 µm ["B. erythrocarpum complex" pro parte] | 22 |
| 22 | Mid laminal cells 14–18 µm wide, thin-walled; tubers often produced on short rhizoids arising in axils of lower leaves | Bryum rubens |
| 22' | Mid laminal cells c. 10 µm wide, mostly firm-walled; tubers not on short axillary rhizoids | 23 |
| 23 | Tubers mostly >200 µm diam., with cell walls protruding | 24 |
| 23' | Tubers mostly <200 µm diam., with cell walls not protruding | 25 |
| 24 | Tubers brown or red-brown (by reflected light), mostly 260–350 µm diam.; mid laminal cells firm-walled | Bryum duriusculum |
| 24' | Tubers bright yellow-brown (by reflected light), mostly 210–250 µm diam.; mid laminal cells thin-walled | Bryum tenuisetum |
| 25 | Tubers bright orange and attached to purple rhizoids (by reflected light) | Bryum ruderale |
| 25' | Tubers red-brown and attached to brown or red-brown rhizoids (by reflected light) | Bryum radiculosum |
| 26 | Plants serially comose; laminal cells adjacent to costa at mid leaf obliquely oriented; costa concolourous with lamina, short-excurrent | Bryum crassum |
| 26' | Plants not serially comose; laminal cells adjacent to costa at mid leaf not obliquely oriented; costa red at base and long-excurrent | Bryum caespiticium |
| 27 | Leaves comose (often serially) | 28 |
| 27' | Leaves evenly spaced on stem | 29 |
| 28 | Leaves little altered when dry; leaf border weak and often obscured by revolute margins; capsules oblong-cylindric, ≤2.7 mm long; spores 8–10 µm | Bryum crassum |
| 28' | Leaves much altered (often spiralled around leaf axis) when dry; leaf border obvious; capsules narrowly cylindric, usually >4 mm long; spores >10 µm, often variable in single capsule | Rosulabryum subtomentosum (small forms) |
| 29 | Leaves much altered when dry, decurrent at base (especially obvious in sterile plants); upper laminal cells moderately thick-walled, not obliquely oriented; synoicous in N.Z. | Bryum pseudotriquetrum |
| 29' | Leaves scarcely altered when dry, not decurrent; upper laminal cells strongly incrassate, mostly obliquely oriented; dioicous (but males inconspicuous) | Bryum laevigatum |
| 30 | Exostome teeth with lamellae joined by longitudinal and oblique thickenings; peristome appearing single under low magnification (endostome adhering to teeth and often fugacious) | Bryum algovicum var. rutheanum |
| 30' | Exostome teeth lacking longitudinal thickenings between lamellae; peristome obviously double under low magnification (endostome not adherent) | 31 |
| 31 | Costa excurrent to form a slender awn; leaves moderately to strongly comose | 32 |
| 31' | Costa short-excurrent to form a mucro or percurrent; leaves not or weakly comose | 34 |
| 32 | Dioicous; leaves indistinctly bordered, the border not extending to apex (and usually obscure at mid leaf shoulder); upper laminal cells firm-walled; costa yellow-brown throughout; cilia elongate, nodose; spores 15–21 µm | Rosulabryum campylothecium |
| 32' | Synoicous; leaves distinctly bordered to apex; upper laminal cells thin-walled; costa red at base or nearly throughout; cilia rudimentary; spores always >20 µm | 33 |
| 33 | Plants small and catkin-like, with stems mostly <7 mm; leaves not decurrent, those of comae <2.5 mm; setae to c. 15 mm; capsules c. 2 mm long; exostome teeth with c. 12–14 adaxial lamellae | Bryum tenuidens |
| 33' | Plants not catkin-like, stems mostly >15 mm; leaves weakly decurrent, those of comae mostly >3 mm; setae mostly >35 mm; capsules c. 4 mm; exostome teeth with c. 25 or more adaxial lamellae | Bryum amblyodon |
| 34 | Capsules horizontal to suberect, obovoid-cylindric; spores 8–12 µm; dioicous; setae 7–10 mm | Bryum preissianum |
| 34' | Capsules nodding or inclined, not obovoid-cylindric; spores larger, mostly >20 µm (except in some populations of B. clavatum); sexuality various; setae nearly always ≥15 mm | 35 |
| 35 | Neck c. ⅓ the total capsule length; capsules ≤2.5 mm, narrowly pyriform, not curved, strongly pendent | Bryum pallescens |
| 35' | Neck c. ½ the total capsule length; capsules nearly always 3–5 mm long, clavate or narrowly pyriform, curved or straight, inclined to pendent | 36 |
| 36 | Capsules asymmetric and curved, clavate, (2.5–)3.5–5 mm long; upper laminal cells firm-walled, c. 27–45 × 9–12 µm; dioicous; tubers sometimes present, red-brown, irregular, mostly 75–150 µm in greater diam.; plants widespread at elevations below c. 1220 m | Bryum clavatum |
| 36' | Capsules symmetric or nearly so, narrowly pyriform, 3–5 mm long; upper laminal cells thin-walled, 69–90 × 24–27 µm; synoicous; tubers not known; plants known from few localities from c. 1100–1800 m | Bryum mucronatum |
Biostatus
Indigenous (Non-endemic)
| Category | Number |
|---|---|
| Indigenous (Endemic) | 2 |
| Indigenous (Non-endemic) | 25 |
| Exotic: Fully Naturalised | 3 |
| Total | 30 |
Excluded Taxa
The genus Anomobryum Schimp. was recorded from N.Z. by Dixon (1926) to accommodate the species treated here as Bryum harriottii. The placement of this species in Bryum agrees with both Ochi (1992) and Spence & Ramsay (2006).
The genus Brachymenium Schwägr. has been used by several authors (Dixon 1926; Sainsbury 1955b; Ochi 1970) to accommodate the Australasian species treated here as Bryum preissianum. The rationale for placing this species in the genus where it was originally described is provided under the species discussion, below. Brachymenium is represented in Australia by two species (Spence & Ramsay 2006), neither of which occurs in N.Z.
The genus Gemmabryum was described by Spence & Ramsay (2005) as a segregate of Bryum. Those authors (2006, p. 287) characterised species of Gemmabryum as having "Bryum-like areolation" and stems that are bud-like or elongate and non-julaceous, and leaves mostly with weak costae that do not reach the leaf apex. Contrary to this characterisation, many of the species they illustrate have excurrent costae. They recognised three distinct types of gemmae in Gemmabryum and placed 25 Australian species (of which 12 are accepted from N.Z. in Bryum, sometimes under differing epithets) in this genus. The genus Gemmabryum is not accepted here because of its apparent heterogeneity and my inability to recognise it in the field.
The genus Imbribryum was described by Pedersen (2005) with Bryum alpinum With. as its type. Spence & Ramsay (2013) recently transferred five Australasian species to this genus, which is neither accepted nor considered further here.
The genus Leptobryum (Bruch & Schimp.) Wilson was treated in the Bryaceae by Sainsbury (1955a). In this Flora it is treated in the Meesiaceae, following Goffinet et al. (2009).
The genus Leptostomum R.Br. was treated in the Bryaceae by Sainsbury (1955a), but its placement there has long been controversial (Ramsay 2006). In this Flora it is treated in the monotypic family Leptostomataceae in the Bryales, in accordance with its treatment by both Ramsay (2006) and Goffinet et al. (2009).
The genus Mielichhoferia Nees & Hornsch. was treated in the Bryaceae by Sainsbury (1955a). Shaw & Crum (1984) examined the peristome structure of North American species placed in Mielichhoferia, and Shaw (1985) subsequently resurrected the genus Schizymenium Harv. for species excluded from a narrowly-defined Mielichhoferia by their monoicous sexuality, and by peristomes entirely endostomal in nature or double and with unornamented teeth. The relevant N.Z. species are referred to Schizymenium, and are discussed in this Flora in the family Mniaceae, following the family placement of Goffinet et al. (2009).
The genus Ochiobryum was described by Spence & Ramsay (2005), based on Bryum blandum Hook.f. & Wilson, a species with a Campbell I. type. This species occurs in both Australia and N.Z. and has been retained in Bryum by Australasian authors since its original description. However, as pointed out by Spence & Ramsay (2006), it is highly anomalous there and deserves generic isolation. Spence & Ramsay (2006, p. 320) characterised Ochiobryum as having "frequently complanate foliation, shiny pink to red colouration, elongate, pohlioid laminal areolation with a distinct border, and inclined to pendulous pyriforme capsules with complete peristomes and small spores". The genus Ochiobryum is accepted here but is placed in the family Mniaceae, in accordance with Goffinet et al. (2009).
The genus Orthodontium Schwägr. was treated in the Bryaceae by Sainsbury (1955a). In this Flora it is treated in the Orthodontiaceae, following its placement by Goffinet et al. (2009).
The genus Pohlia Hedw. was treated in the Bryaceae by Sainsbury (1955a). In this Flora it is treated in the Mniaceae, following Goffinet et al. (2009) and other recent authors (e.g., Smith 2004).
The genus Ptychostomum Hornsch. was resurrected by Spence (2005) and Spence & Ramsay (2006) subsequently placed five Australian species in this genus, two of them accepted from N.Z. They (p. 323) characterised members of Ptychostomum as having "rhodobryoid laminal areolation, peristome reduction associated with nutant capsules, and a lack of asexual propagules other than occasional axillary filiform gemmae". The genus Ptychostomum is not accepted here.
Bibliography
Brotherus, V.F. 1924: Musci (Laubmoose). II. Spezieller Teil. In: Engler, A. (ed.) Die natürlichen Pflanzenfamilien. Edition 2. Bd 10. Engelmann, Leipzig. 143–478.
Cox, C.J.; Hedderson, T.A.J. 2003: Phylogenetic relationships within the moss family Bryaceae based on chloroplast DNA evidence. Journal of Bryology 25: 31–40.
Crum, H.A.; Anderson, L.E. 1981: Mosses of Eastern North America. Columbia University Press, New York.
Dixon, H.N. 1926: Studies in the bryology of New Zealand, with special reference to the herbarium of Robert Brown. Part IV. Bulletin, New Zealand Institute 3(4): 153–238.
Fife, A.J. 2015: Bryaceae. In: Heenan, P.B.; Breitwieser, I.; Wilton, A.D. (ed.) Flora of New Zealand — Mosses. Fascicle 19. Manaaki Whenua Press, Lincoln.
Goffinet, B.; Buck, W.R.; Shaw, A.J. 2009: Morphology, anatomy, and classification of the Bryophyta. In: Goffinet, B.; Shaw, A.J. (ed.) Bryophyte Biology. Edition 2. Cambridge University Press, Cambridge. 55–138.
Ochi, H. 1970: A revision of the subfamily Bryoideae in Australia, Tasmania, New Zealand, and the adjacent islands. Journal of the Faculty of Education, Tottori University. Natural Science 21(1): 7–67.
Ochi, H. 1992: A revised infrageneric classification of the genus Bryum and related genera (Bryaceae, Musci). Bryobrothera 1: 231–244.
Pedersen, N. 2005: Validation of Imbribryum (Bryaceae). Bryologist 108: 449.
Pedersen, N.; Holyoak, D.T.; Newton, A.E. 2007: Systematics and morphological evolution within the moss family Bryaceae: A comparison between parsimony and Bayesian methods for reconstruction of ancestral character states. Molecular Phylogenetics and Evolution 43: 891–907.
Ramsay, H.P. 2006: Leptostomataceae. In: McCarthy, P.M. (ed.) Flora of Australia. Vol. 51 Mosses 1. ABRS, Canberra. 349–353.
Reichenbach, H.G.L. 1829 ("1828"): Botanik für Damen, Künstler und Freunde der Pflanzenwelt überhaupt, enthaltend eine Darstellung des Pflanzenreichs in seiner Metamorphose. Cnobloch, Leipzig.
Sainsbury, G.O.K. 1955a: A handbook of the New Zealand mosses. Bulletin of the Royal Society of New Zealand 5: 1–490.
Sainsbury, G.O.K. 1955b: Notes on Tasmanian mosses from Rodway's herbarium: IV. Papers and Proceedings of the Royal Society of Tasmania 89: 21–35.
Shaw, A.J. 1985: Nomenclatural changes in the Bryaceae subfamily Mielichhoferioideae. Bryologist 88: 28–30.
Shaw, A.J.; Crum, H.A. 1984: Peristome homology in Mielichhoferia and a taxonomic account of the North American species. Journal of the Hattori Botanical Laboratory 57: 363–381.
Smith, A.J.E. 1978: The Moss Flora of Britain and Ireland. Cambridge University Press, Cambridge.
Smith, A.J.E. 2004: The Moss Flora of Britain and Ireland. Edition 2. Cambridge University Press, Cambridge.
Spence, J.R. 1996: Rosulabryum genus novum (Bryaceae). Bryologist 99: 221–225.
Spence, J.R. 2005: New genera and combinations in Bryaceae (Bryales, Musci) for North America. Phytologia 87: 15–28.
Spence, J.R.; Ramsay, H.P. 1999: Three new species of Rosulabryum (Bryopsida, Bryaceae) from Australia. Telopea 8: 325–335.
Spence, J.R.; Ramsay, H.P. 2005: New genera and combinations in the Bryaceae (Bryales, Musci) for Australia. Phytologia 87: 61–72.
Spence, J.R.; Ramsay, H.P. 2006: Bryaceae. In: McCarthy, P.M. (ed.) Flora of Australia. Vol. 51 Mosses 1. ABRS, Canberra. 274–348.
Spence, J.R.; Ramsay, H.P. 2013: Additions to and nomenclatural changes in the Bryaceae (Bryopsida) in Australia. Telopea 15: 143–148.
Vitt, D.H. 1984: Classification of the Bryospida. In: Schuster, R.M. New Manual of Bryology. Hattori Botanical Laboratory, Nichinan. 696–759.