Class
Family
Genus
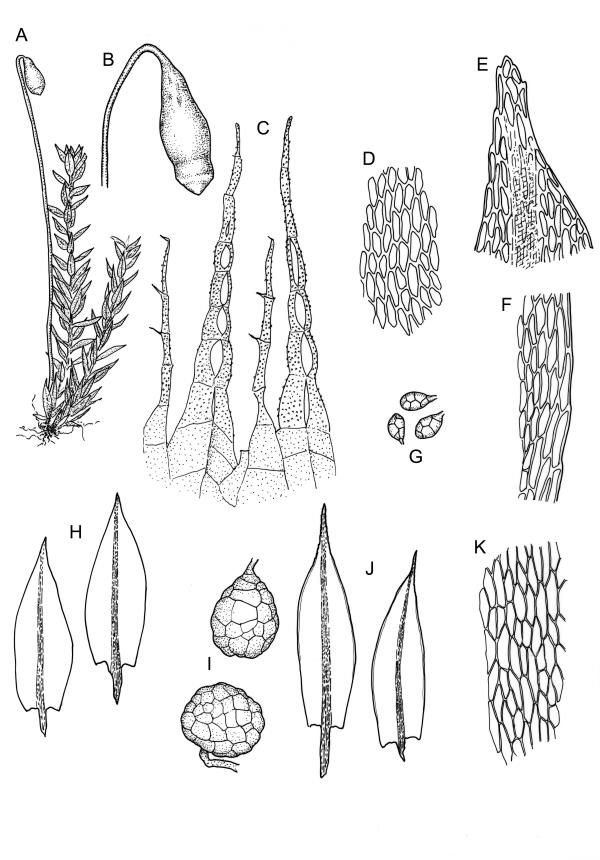
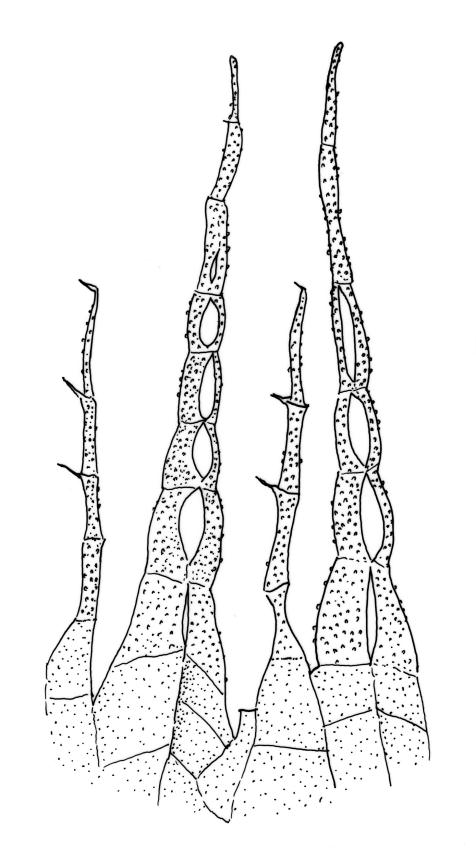
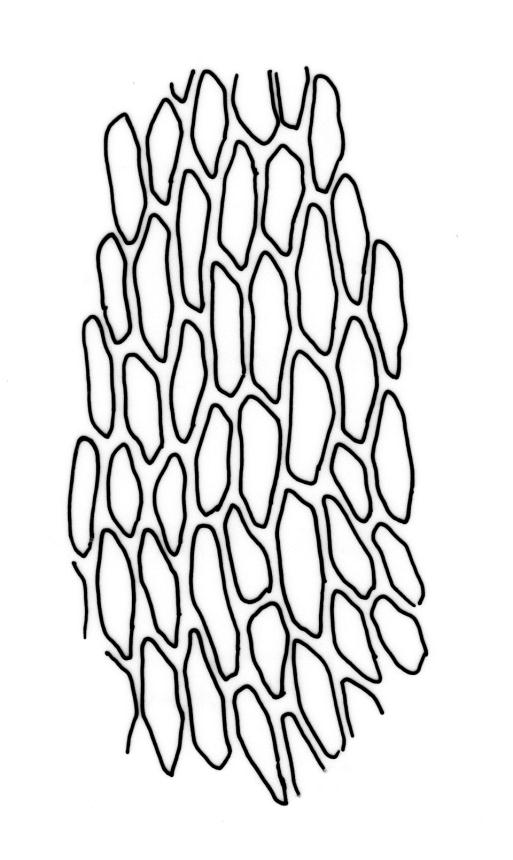
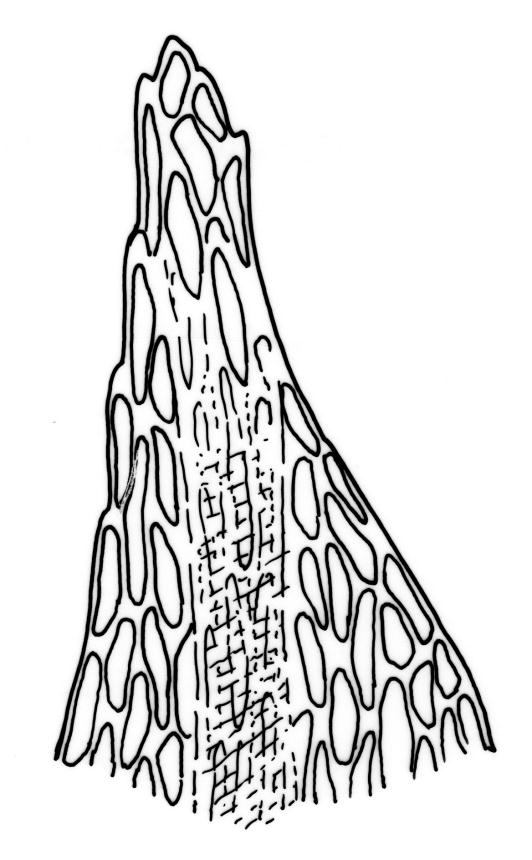
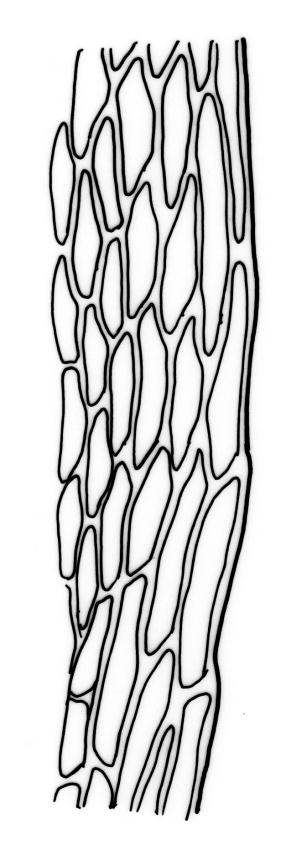
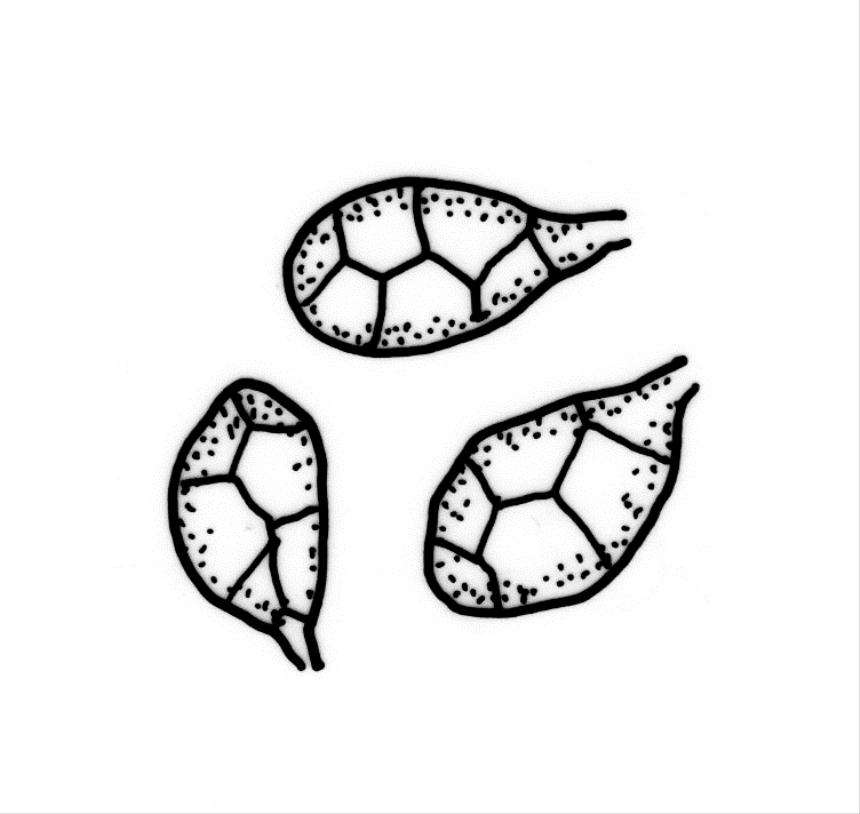
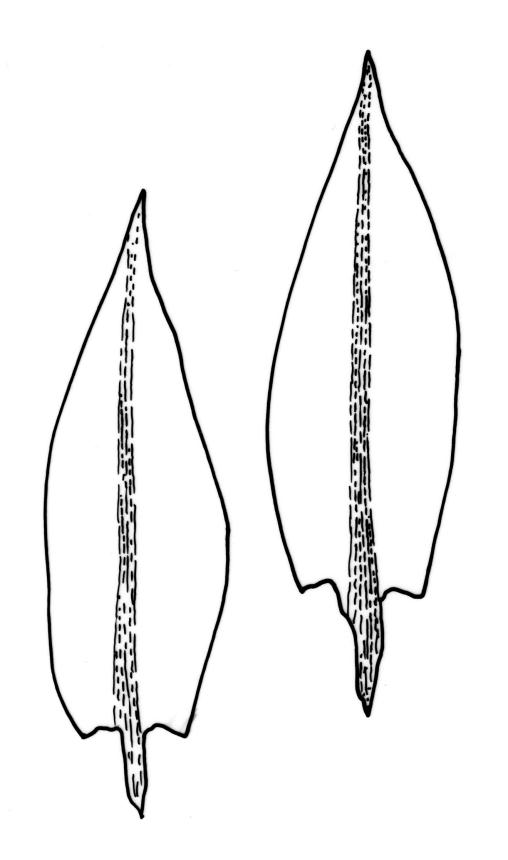
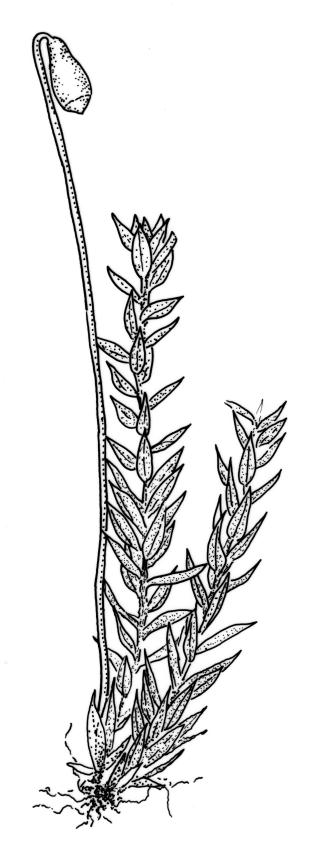
Stems to 15 mm, beset below with red-brown rhizoids. Leaves evenly spaced on stem, erect-spreading when moist, becoming more erect and narrowed when dry, ovate- or elliptic-lanceolate, acute, denticulate above, weakly bordered and ± recurved at margins, not decurrent, c. 1.0–1.8 × 0.5–0.7 mm, with lamina forming 0.85 to ± entire leaf length; mid laminal cells firm-walled (rarely ± thin-walled), (24–)30–48(–75) × 10–12 µm, and c. 3–4:1; marginal cells elongate in 2–4(–5) rows, forming a weak or rarely distinct border; basal cells usually not pigmented. Costa stout, short-excurrent. Tubers pyriform or ellipsoid, (45–)60–105(–140) × 45–60 µm, red-brown, mostly 2 cells wide, with cell walls not protruding.
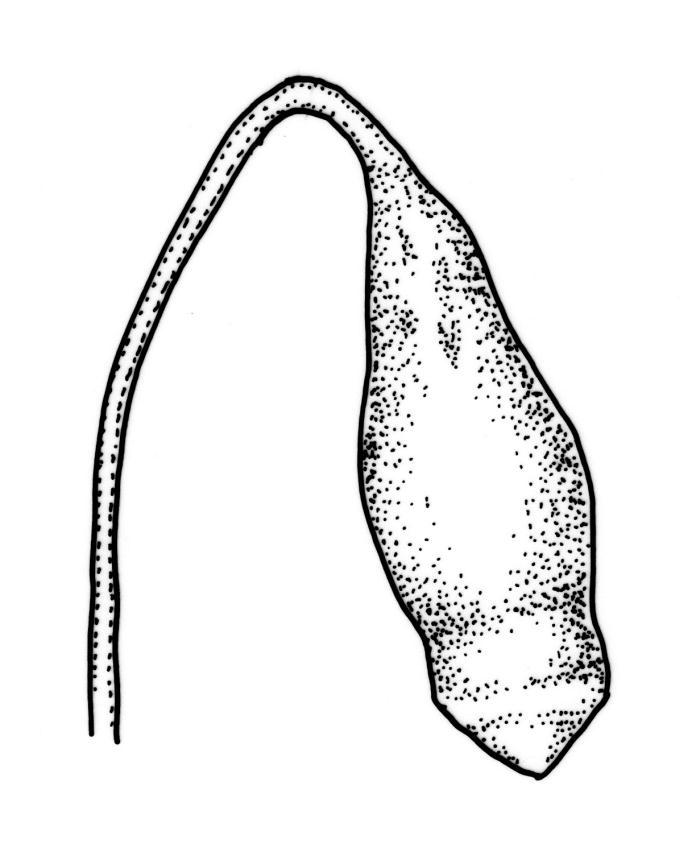
Dioicous. Perichaetia sometimes 2–3 on one plant, appearing to be in branch axils due to innovation, with narrowly lanceolate leaves, c. 2.2 mm. Perigonia terminal or on short lateral branches. Setae to c. 20 mm, red; capsules distinctly constricted below mouth when dry, 1.5–2.0(–3.0) mm long; operculum low-conic, not apiculate. Exostome as per B. erythrocarpum complex; endostome with single appendiculate cilia (rarely 2–3). Spores 14–18(–20) µm.
Crundwell & Nyholm 1964, fig. 5 a–l; Smith 2004, fig. 190, 1–3; Porley 2008, p. 64. Seppelt 2004, fig. 43.
The red costal base cited by Crundwell & Nyholm (1964) for members of the "B. erythrocarpum complex" in Europe is not a constant feature of N.Z. B. sauteri. In most populations leaves on older stem portions have some degree of secondary pigmentation, while those on younger stems have pale or brown lower costae. Crundwell & Nyholm (1964) suggest that both monoicous and dioicous races of B. sauteri occur in Europe, while Smith (2004) stated that in Europe/Britain it is "possible that there are two taxa, one oceanic and dioicous and the other continental and synoicous". N.Z. material appears to be consistently dioicous.
Some sterile populations can be difficult to distinguish from other species, especially Bryum dichotomum, which also has uniformly distributed and weakly bordered leaves, stout, ± excurrent costae, and firm-walled laminal cells. In B. dichotomum the leaves are widest at or near the leaf base and the margins plane while in B. sauteri the leaves are widest at or above the middle and the margins usually ± recurved. Axillary bulbils are usually present in B. dichotomum, but not in B. sauteri.
Distinctions between B. sauteri (and other members of the "B. erythrocarpum complex") and B. clavatum are discussed under the latter species.
K (sterile material); NI: N Auckland, including offshore islands (Bay of Islands, Ōkaihau, Manukau, and sterile material from Kaipara Harbour, Kawerua, TK), S Auckland (Tuakau and sterile material from Whale I.), Gisborne (sterile material from Te Waipuhake Stream),Wellington (Tiritea River, Mana I., Hutt Valley, Wiltons Bush, and sterile material from Mt Ruapehu, Somes I.); SI: Nelson (Nine Mile Beach, and sterile material from Stephens I., Kōhaihai Bluff, Perpendicular Point), Marlborough (D'Urville I., and sterile material from Puhi Puhi River, Kaikōura), Canterbury (Port Hills, summit of Arthur’s Pass, and sterile material from Banks Peninsula, Broad Stream, Erewhon, Hooker Valley), Otago (Signal Hill, Berwick, Sandymount); St (sterile material only); Ch (sterile material only); C (sterile material only); M (sterile material only). Localities given are for fertile material unless specified.
Apparently bipolar. Reported from Tasmania and mainland Australia by Spence & Ramsay (2006), from Britain and scattered European localities by Crundwell & Nyholm (1964), as well as North America, south-east Asia, New Guinea, Lord Howe I., and Norfolk I. by Spence & Ramsay (2006).
Coastal or inland, mostly on vertical soil banks and often associated with weak seepage. Occasionally on bare soil, or over rock (pumice, papa, conglomerate, etc) and nearly always occurring in ± basic (nutrient rich?) situations. Frequently associated species include Bryum dichotomum, Calyptrochaeta apiculata, Hennediella macrophylla, Philonotis tenuis, Pseudocrossidium crinitum, Tridontium tasmanicum, Weissia austrocrispa, and W. controversa. Where influenced by coastal spray, occurring from sea level to c. 325 m, but extending to c. 900 m (and rarely to c. 1300 m at Broad Stream, Canterbury L.D.) in inland situations.
Bryum sauteri is one of six N.Z. members of the "B. erythrocarpum complex" sensu Crundwell & Nyholm (1964). The complex is discussed in more detail under B. duriusculum. With the exception of B. duriusculum, it is the most commonly collected member of the complex in N.Z.
While the ellipsoid to pyriform tubers appear to be constantly present, they can sometimes be sparse. The highly characteristic tubers make this species unmistakeable; the capsules when present are characteristically constricted below the mouth. In a small fraction of populations the laminal cells are relatively thin-walled; such variation appears to be developmental, with thinner cell walls restricted to younger plants.
Some material from Chatham I. (A.J. Fife 11282, CHR 514610) and Perpendicular Point, Nelson L.D. (A.J. Fife 5018, CHR 103708) is unusual in having a very strong leaf borders (4–5 cell rows wide at mid leaf) and plane, denticulate leaf margins.