Class
Family
Genus
Bryum microerythrocarpum sensuScott & Stone (1976); sensu Beever et al. (1992)
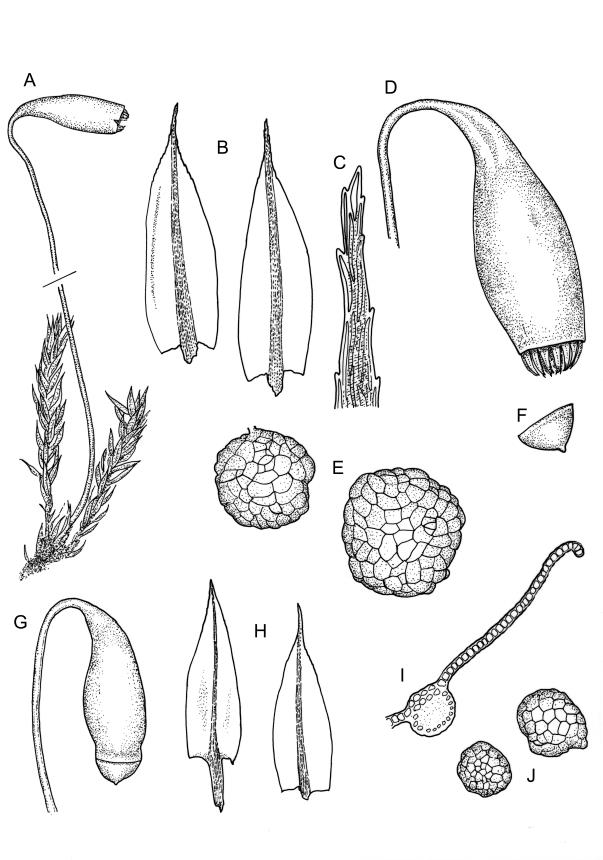
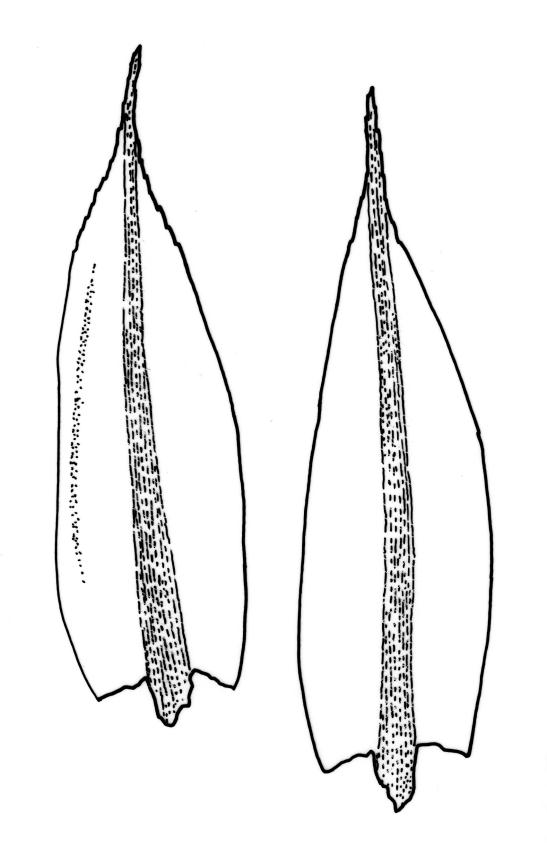
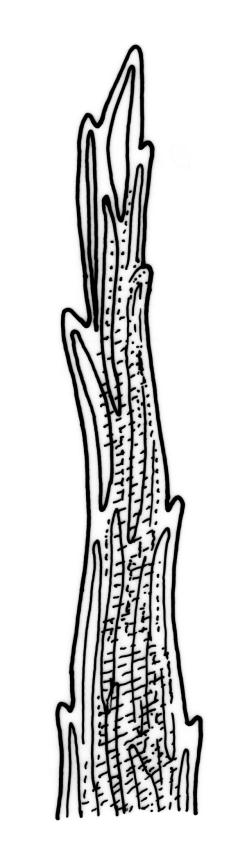
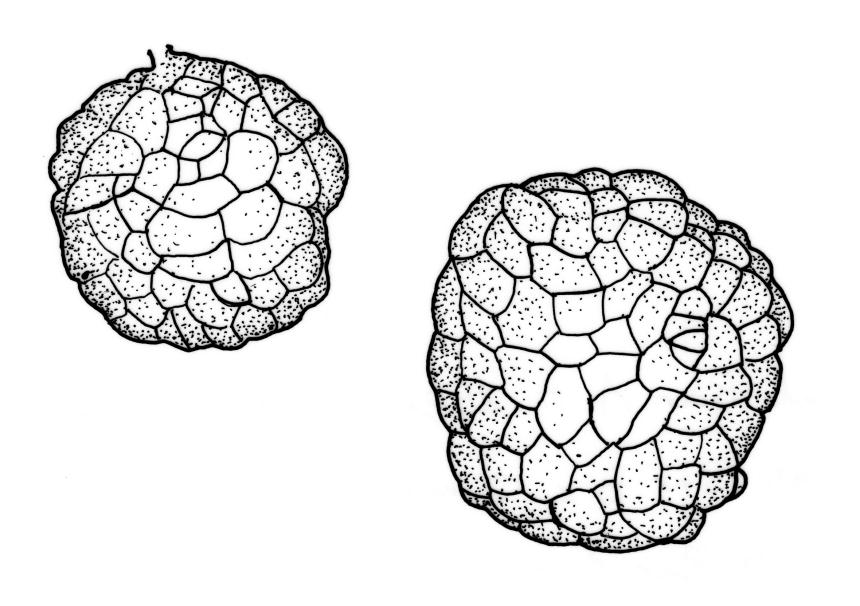
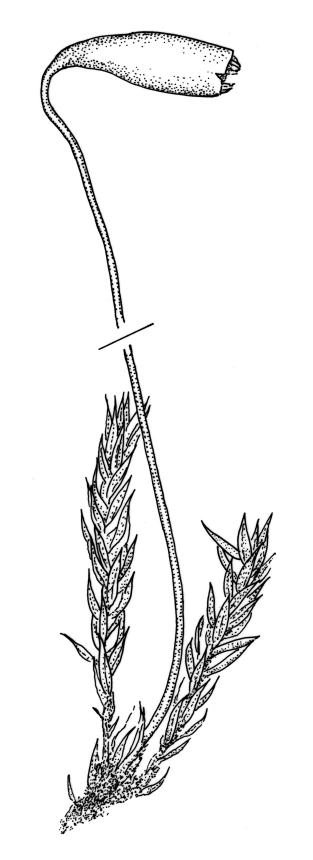
Stems to c. 15 mm, beset below with red-brown rhizoids. Leaves evenly spaced on stem, erect-spreading when moist, becoming more erect and narrowed when dry, ovate- or elliptic-lanceolate, acute, denticulate above, weakly bordered and ± recurved at margins, neither plicate nor decurrent, 1.0–1.6 × 0.3–0.5 mm, with lamina forming 0.85–0.9 of entire length; mid laminal cells firm-walled, c. 30–60 × 9–12 µm; marginal cells elongate in 1–2 rows and forming a very weak border. Costa stout and excurrent to form a denticulate awn. Tubers ± spherical, oblong, or somewhat irregular, (225–)260–350 µm diam., brown or red-brown, c. 10 cells across, with cells strongly or scarcely protruding (a few tubers sometimes on short rhizoids in leaf axils).
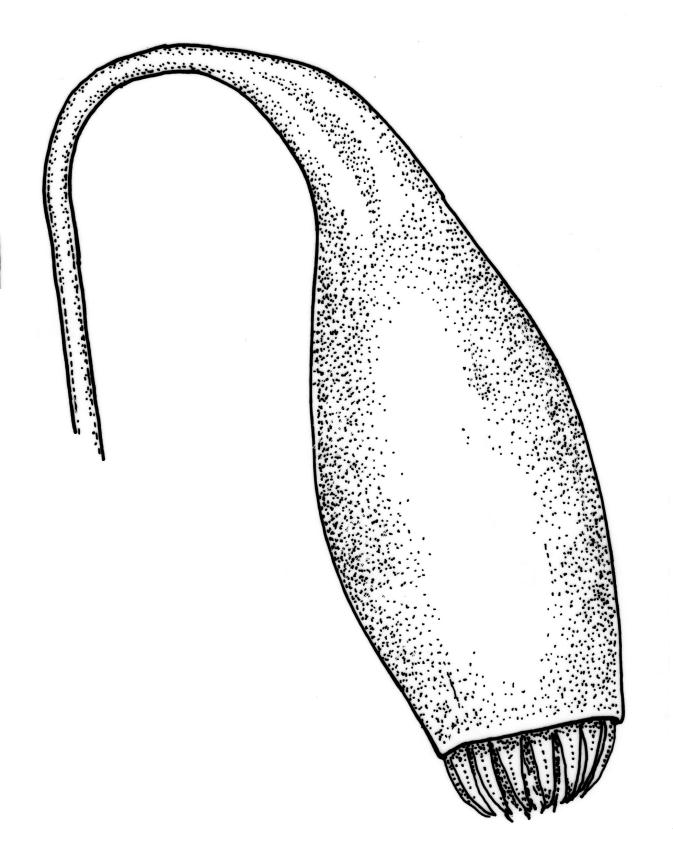
Dioicous. Perichaetia 1–3 per shoot, often appearing axillary due to innovation, with leaves narrowly lanceolate or deltoid. Perigonia not seen. Setae c. 20–45 mm, red; capsules weakly constricted below mouth when dry, (1.8–)2.0–3.5 mm long; operculum conic and apiculate. Exostome teeth red-brown below, pale and finely baculate above; endostome with 2–3 appendiculate cilia. Spores 9–12 µm.
Beever et al. 1992, fig. 40 h (as B. microerythrocarpum).
Bryum duriusculum has upper leaf margins more denticulate and costae and basal laminal cells more consistently suffused with red pigment than B. sauteri, which is also widespread in N.Z.
K (sterile material only); NI: N Auckland, S Auckland, Gisborne (Lake Waikaremoana, Mt Manuoha, Ōhinepākā), Hawke's Bay (Wairoa, Matamau), Wellington (Ruahine Range, Akatarawa); SI: Nelson (Farewell Spit), Marlborough (Kenepuru and sterile material from Blue Duck Reserve), Canterbury (Arthurs Pass), Westland (Ōtira, Lake Brunner, and sterile material from Hokitika); Otago (Dunedin, Pine Hill), Southland (Mossburn, Puysegur Point); St (fertile and sterile material); Ch (sterile material only); C (fertile and sterile material). Localities given are for fertile material unless specified.
Probably widespread outside N.Z. but no attempt has been made to summarise the extra-N.Z. distribution because the relationship to B. microerythrocarpum Müll.Hal. & Kindb. needs monographic study for clarification.
On sandy or humic soil, sometimes in bogs, often beneath manuka (Leptospermum scoparium) and sometimes on charred soil; from low elevation to at least 1130 m (on Mt Ruapehu, Wellington L.D.). Bryum duriusculum occurs on acidic and often coarsely grained soil. Rarely epiphytic on accumulated humus. Very often growing mixed with Ceratodon purpureus, as well as Campylopus introflexus, Funaria hygrometrica, and Leptobryum pyriforme.
Bryum duriusculum is the most widespread and commonly collected representative of the "B. erythrocarpum complex" (discussed below) in N.Z. Much of the N.Z. material referred by previous workers to B. chrysoneuron Müll.Hal. (see "Excluded Taxa" above) is referable here. Bryum duriusculum is the earliest typifiable name for N.Z. material of the complex with large (c. 260–350 µm), spherical, red-brown tubers.
Wilson's citations of B. duriusculum syntypes are difficult to interpret. However, in Wilson's herbarium, his handwritten description refers clearly to Lyall 232 and Colenso 67b. Neither of these collections are attached to the same sheet as the description, however material of Lyall 232, annotated by Wilson, is present in the Hooker herbarium. This material bears both capsules and the rhizoidal tubers necessary for the confident determination of a species in the B. erythrocarpum complex, and is therefore designated the lectotype. Bryum duriusculum is probably an earlier name for the widespread B. microerythrocarpum Müll.Hal. & Kindb., which is discussed under "Excluded Taxa" above.
Material from the Pongakawa River (S Auckland L.D.; K.W. Allison 2466, CHR 490334) has very sparse tubers with morphology typical of the species, but its overall habit is laxer than other collections. It grew immersed or nearly so in a permanent spring and is referred here with reservation.
Bryum duriusculum is considered here to be a member of the "B. erythrocarpum complex" sensu Crundwell & Nyholm (1964). The six species belonging to this complex documented from N.Z. are: B. duriusculum, B. radiculosum, B. rubens, B. ruderale, B. sauteri, and B. tenuisetum. The delineation of these species is largely reliant on tuber morphology. Crundwell & Nyholm’s (1964) revision of the European members of the complex remains useful, even in N.Z. Considerable emphasis was placed by Crundwell & Nyholm on whether or not the individual cells of water-mounted tubers protrude beyond the general outline of the tuber, and tuber shape, diameter, and pigmentation. An excellent series of photographs of tubers of British members of the complex is provided by Porley 2008, p. 62 et seq.).
N.Z. collections with well-developed tubers can be named confidently and the distinctions between the six accepted species are convincing. However, the tubers can be sparse or rarely absent and hence not all material can be named to species level. Members of the complex have a recognisable facies deriving from their rather small shoots, non-comose, erect-spreading, weakly bordered, and denticulate leaves, usually firm-walled laminal cells, stout and short-excurrent costae, and pendent, pyriform capsules. Because of their distinctive facies, a supplementary description applicable to all members of the complex is given here.
"Bryum erythrocarpum complex" sensu Crundwell & Nyholm
Plants bright, yellow-green or bronze, forming turves. Stems ± red below or throughout, branching by innovation, beset with red-brown, purple or yellow-brown, coarsely papillose rhizoids, in cross-section with firm-walled cortical cells and a weak central strand. Leaves not or weakly crowded at stem apices, erect-spreading, becoming more erect and narrowed when dry, ovate- or elliptic-lanceolate, acute, <1–c. 2.5 mm, concave, denticulate above (rarely only near apex), weakly bordered, mostly ± recurved, not decurrent; upper laminal cells narrowly hexagonal, firm-walled or rarely thin-walled, longer and more oblong below; marginal cells longer, forming an ill-defined or rarely distinct border 1–4(–5) cells wide at mid leaf; basal cells short, ± oblong, with or without secondary pigments. Costa stout or rarely thin, variably coloured (red, brown, or pale below), short- or long-excurrent, usually forming a denticulate awn. Brood bodies (rhizoidal tubers) nearly constantly present.
Dioicous and infrequently fruiting. Perichaetia 1-3 per shoot, often appearing axillary due to innovation, with leaves narrowly lanceolate or deltoid. Perigonia terminal or on short lateral branches. Setae cygneous just below capsule; capsules pendent, narrowly pyriform, constricted below mouth when dry, with a neck c. ⅓ the total length and only weakly wrinkled; operculum lowly conic, mostly apiculate (non-apiculate in B. radiculosum). Exostome teeth red-brown below, pale and coarsely baculate above; endostome with fenestrate segments and well-developed, appendiculate cilia. Spores < 20 µm diam.
The late J.T. Linzey took a special interest in the "B. erythrocarpum complex" in N.Z. His results were never published but his still useful manuscript is preserved at AK and is referred to here.
Confusion occurs between the members of the "B. erythrocarpum complex" and B. clavatum, particularly with sterile material. The toothed leaf apices and the weaker leaf borders in the complex provide some distinction, as do the short-oblong basal leaf cells. When fertile, the pendent and symmetric capsules in species of the complex contrast with the usually inclined and curved capsules of B. clavatum. The distinctions are discussed further under B. clavatum.
Linzey (unpublished) recognised, but never formally described, what he considered a distinctive species in this "species complex", which he termed "Bryum ACC" (after the British bryologist Alan C. Crundwell). These plants have red-brown tubers 60-105 µm diam., ± irregular in outline and with individual cells moderately protruding. The bulk of material so-named by Linzey is considered here not to be a member of the complex but to be aberrant B. clavatum (q.v.); it is discussed further under that species.
Numerous collections, mostly sterile and apparently belonging to this complex, cannot be named with confidence. Among the most distinctive are two North Auckland L.D. (CHR 106581 & 106609) and two Nelson L.D. (CHR 406023 & 405538) collections made from marl soil. These four specimens have lanceolate, distinctly bordered, and entire leaves with long-excurrent costae (to ⅓ the total leaf length), and abundant, irregular, brown tubers 90-180 µm diameter with cells moderately to strongly protruding. Further study may show this material to be worthy of description. Other aberrant and indeterminable material has been seen from Mt Ruapehu, Wellington L.D. (J.E. Beever 31-01, CHR 405923); Nile River, Nelson L.D. (A.J. Fife 6015, CHR 405539); and Raramai Tunnel, Marlborough L.D. (J. Lewinsky 1087, CHR 348510).
Ochi's (1992, p. 235) concept of the subsect. Apalodictyon roughly corresponds to the "B. erythrocarpum complex" as treated here.