Class
Classification
Subordinate Taxa
Nomenclature
Scientific Name:
Rhizogoniaceae Broth., Nat. Pflanzenfam. [Engler & Prantl] 1(3), 614 (1904)
Type Taxon:
Description
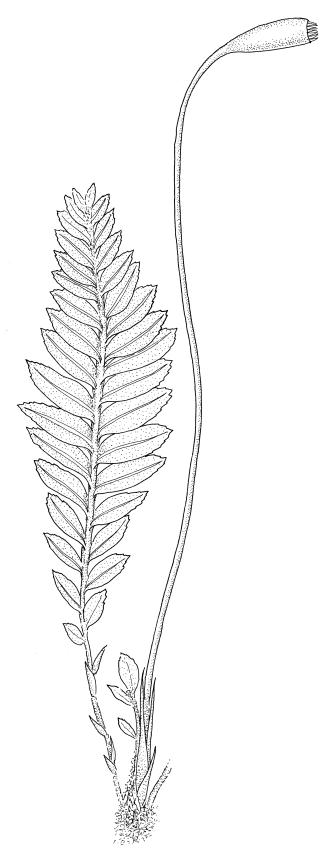
Plants small to robust, epiphytic or terrestrial. Stems erect, simple or much branched, in cross-section angular in outline, with a central strand. Shoots often complanate. Leaves either uniform or dimorphic, distichous, tristichous, or inserted in many rows; those of branches similar or rarely differentiated from stem leaves, often bordered, mostly sharply toothed (with teeth either single or paired), less often denticulate or entire; laminal cells oblong, rounded-hexagonal or rarely oblong-hexagonal, usually thick-walled and compact, rarely lax (in Goniobryum), smooth; alar cells not differentiated. Costa strong, ending below the apex to excurrent.
Perichaetia mostly basal or lateral, occasionally terminal (in Calomnion and Cryptogonium). Setae single or rarely aggregated, mostly elongate; capsules horizontal, inclined or erect, exserted or rarely immersed (in Cryptogonium), oblong-cylindric to cylindric; peristome mostly double and well-developed, rarely absent (in Calomnion); annulus differentiated; operculum conic or stoutly rostrate. Calyptra cucullate and smooth.
Taxonomy
Perichaetia in this family are usually borne at the stem base or low on the stem. However, the perichaetia in Calomnion and Cryptogonium (the former gymnostomous and often placed in its own family) are terminal, and in Cryptogonium the perichaetia are overtopped by innovations.
The difficulty of circumscribing the Rhizogoniaceae and its allies is reflected by its complicated history. Allied (or purportedly allied) genera for many decades have been moved between the present family, the Aulacomniaceae, the Calomniaceae, and, more recently, the Orthodontiaceae.
The Rhizogoniaceae were proposed by Brotherus (1901–1909); in his subsequent (Brotherus 1924) family outline he recognised seven genera occurring mostly in tropical to temperate parts of the southern hemisphere and placed the family in the Bryales ("Eubryales"). Brotherus (1924) recognised four sections within his broadly defined Rhizogonium. Dixon (1926), in his discussion for N.Z., accepted the four N.Z. genera (Cryptopodium, Goniobryum, and Rhizogonium, plus Hymenodon) included there by Brotherus, and added Leptotheca and Aulacomnium. Sainsbury (1955), without comment, followed the placements of Brotherus. In their moss flora of southern Australia, Scott & Stone (1976) also essentially followed a Brotherean concept of the family.
All the treatments above took a broad view of the core genus Rhizogonium Brid. Manuel (1980) took a more restricted view of that genus, resurrecting the long-standing but little-known Pyrrhobryum Mitt. to accommodate species dispersed by Brotherus among three of his four sections of Rhizogonium. The genus Pyrrhobryum (based on the widespread P. spiniforme (Hedw.) Mitt.) has subsequently been accepted by Koponen et al. (1986) in their treatment for the Huon Peninsula, P.N.G. and by Gilmore (2006) in his treatment for Australia.
Churchill & Buck (1982) argued for the transfer of Leptotheca from the Aulacomniaceae to the Rhizogoniaceae. Vitt (1995) presented arguments for the placement of the gymnostomous and problematic Calomnion both "near Rhizogonium" and in the distantly related Tetraphidales, but ultimately retained it in a monogeneric Calomniaceae.
The family Rhizogoniaceae and its allies remain very much in a state of flux. Modern workers have continued to modify the family, anchored by Rhizogonium Brid. (holotype: R. novae-hollandiae (Brid.) Brid.). In a large-scale phylogenetic analysis of "early-diverging" pleurocarpous lineages using both chloroplast and mitochondrial genes, Bell et al. (2007) presented strict consensus trees that demand further reassessment of the Rhizogoniaceae and allies.
Many of the realignments suggested by Bell et al. (2007) are identical to those adopted by Goffinet et al. (2009). Most, albeit not all, of the realignments adopted by the latter are followed here. Those accepted here include the inclusion of Calomnion in the Rhizogoniaceae (rather than in a monogeneric Calomniaceae) and the movement of both Leptotheca and Hymenodon to the Orthodontiaceae.
The assignments, suggested by Bell and co-authors, of two N.Z. taxa (Pyrrhobryum mnioides s.l. and P. bifarium) to the Malesian genus Hymenodontopsis Herzog are rejected here, for reasons outlined under the genus Pyrrhobryum. Given the uncertainty concerning the circumscription and the relationships of Pyrrhobryum, a conservative treatment (a broad interpretation of the genus and its retention in the Rhizogoniaceae) of the genus is adopted here. The family placement of Hymenodontopsis is beyond the scope of this Flora. However, the capsule morphology of the type of the genus, H. steresmannii from Malesia, differs markedly in many ways (capsule shape, its lack of sulci when dry, operculum form, nature of the annulus, and peristome structure) from that of most species of Aulacomnium, a genus to which the analysis of Bell et al. (2007) showed it to be allied.
Key
Key to the genera (N.B.: Hymenodon is included in this key, since it has traditionally been placed in this family and many students will seek it here; in this Flora it is treated in the Orthodontiaceae.)
| 1 | Laminal cells thin-walled and lax, oblong-hexagonal, 3–5:1; autoicous (perigonia scattered amongst perichaetia, brown & bud-like) | Goniobryum |
| 1' | Laminal cells firm- or thick-walled, not lax, variable in shape but <2:1; dioicous (in N.Z. taxa) | 2 |
| 2 | Leaves inserted in three distinct ranks (except at shoot apices), with one rank clearly reduced in size; peristome lacking; protonema persistent | Calomnion |
| 2' | Leaves not in three ranks and lacking a rank of clearly reduced leaves; peristome present, mostly well-developed and double, rarely single; protonema not persistent | 3 |
| 3 | Leaves piliferous, with costa ending well below the base of the hair-point, unbordered, crenulate at margins; laminal cells mammillose; peristome single | Hymenodon [Orthodontiaceae] |
| 3' | Leaves not piliferous, sometimes aristate (due to excurrent costae), bordered or otherwise, serrate to dentate at margins; laminal cells not mammillose; peristome double and well-developed | 4 |
| 4 | Plants very robust, mostly epiphytic on tree ferns; stems c. 50–200 mm; leaves mostly >7 mm and linear-lanceolate; setae <3 mm; capsules immersed and often clustered (to four per perichaetium); perichaetia terminal, but often overtopped by subperichaetial innovations | Cryptopodium |
| 4' | Plants less robust, terrestrial or epiphytic; stems <70 mm (only rarely >50 mm); leaves <6 mm and not linear-lanceolate; setae >10 mm, capsules exserted and single; perichaetia in basal clusters or lateral | 5 |
| 5 | Stem leaves distichous; leaf margins singly toothed; costa smooth abaxially; perichaetia basal | Rhizogonium |
| 5' | Stem leaves not distichous (branch leaves may be); leaf margins doubly toothed; costa toothed abaxially; perichaetia scattered on lower stems or basal (in P. paramattense) | Pyrrhobryum |
Biostatus
Indigenous (Non-endemic)
| Category | Number |
|---|---|
| Indigenous (Endemic) | 1 |
| Indigenous (Non-endemic) | 8 |
| Total | 9 |
Bibliography
Bell, N.E.; Quandt, D.; O'Brien, T.J.; Newton, A.E. 2007: Taxonomy and phylogeny in the earliest diverging pleurocarps: square holes and bifurcating pegs. Bryologist 110: 533–560.
Brotherus, V.F. 1901–1909: Musci (Laubmoose) II Specieller Teil. In: Engler, A.; Prantl, K. (ed.) Die natürlichen Pflanzenfamilien. Teil 1. Abt. 3. Engelmann, Leipzig. 277–1246.
Brotherus, V.F. 1924: Musci (Laubmoose). II. Spezieller Teil. In: Engler, A. (ed.) Die natürlichen Pflanzenfamilien. Edition 2. Bd 10. Engelmann, Leipzig. 143–478.
Churchill, S.P.; Buck, W.R. 1982: A taxonomic investigation of Leptotheca (Rhizogoniaceae). Brittonia 34: 1–11.
Fife, A.J. 2016: Rhizogoniaceae. In: Heenan, P.B.; Breitwieser, I.; Wilton, A.D. (ed.) Flora of New Zealand — Mosses. Fascicle 26. Manaaki Whenua Press, Lincoln.
Gilmore, S.R. 2006: Rhizogoniaceae. In: McCarthy, P.M. (ed.) Flora of Australia. Vol. 51 Mosses 1. ABRS, Canberra. 354–366.
Goffinet, B.; Buck, W.R.; Shaw, A.J. 2009: Morphology, anatomy, and classification of the Bryophyta. In: Goffinet, B.; Shaw, A.J. (ed.) Bryophyte Biology. Edition 2. Cambridge University Press, Cambridge. 55–138.
Koponen, T.; Touw, A.; Norris, D.H. 1986: Bryophyte flora of the Huon Peninsula, Papua New Guinea. XIV. Rhizogoniaceae (Musci). Acta Botanica Fennica 133: 1–24.
Manuel, G. 1980: Miscellanea bryologica II. Classification of Rhizogonium Brid. Penzigiella hookeri Gangulee, and some nomina nuda. Cryptogamie - Bryologie et Lichénologie 1: 67–72.
Sainsbury, G.O.K. 1955: A handbook of the New Zealand mosses. Bulletin of the Royal Society of New Zealand 5: 1–490.
Scott, G.A.M.; Stone, I.G. 1976: The Mosses of Southern Australia. Academic Press, London.
Vitt, D.H. 1995: The genus Calomnion (Bryopsida): taxonomy, phylogeny, and biogeography. Bryologist 98: 338–358.