Class
Family
Genus
Non Bryum platyloma Schwägr., Sp. Musc. Frond. Suppl. 1(2): 116 (1816); nec Bryum capillare var. platyloma (Schwägr.) Schimp., Coroll. Bryol. Eur. 75 (1856); nec Rosulabryum platyloma (Schwägr.) Ochyra, Biodivers. Poland 3: 162 (2003).
Bryum truncorum sensu Sainsbury 1955.
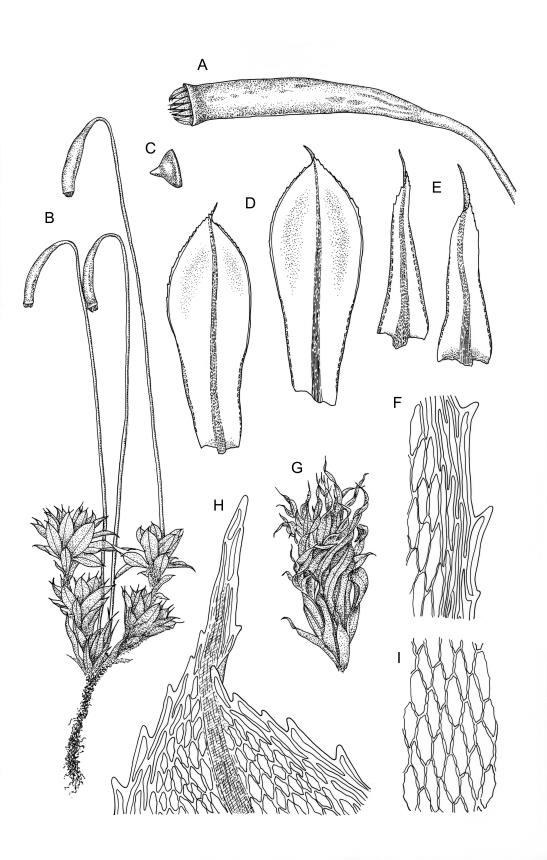
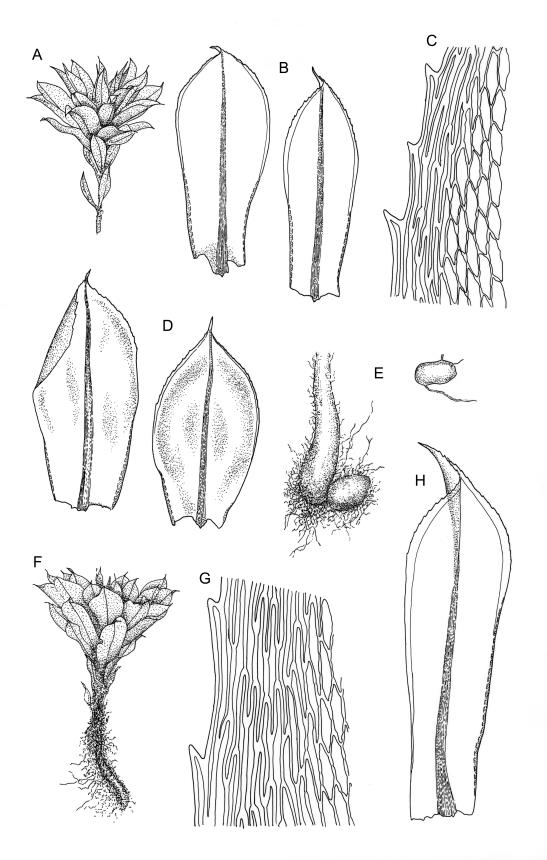
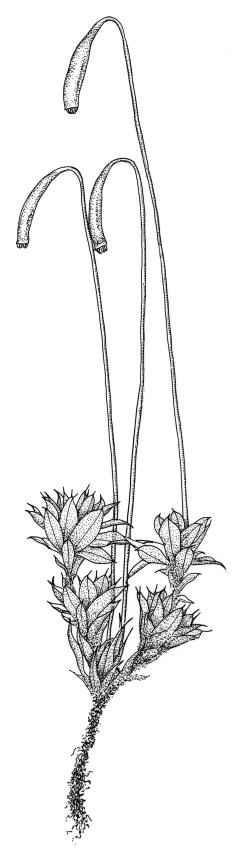
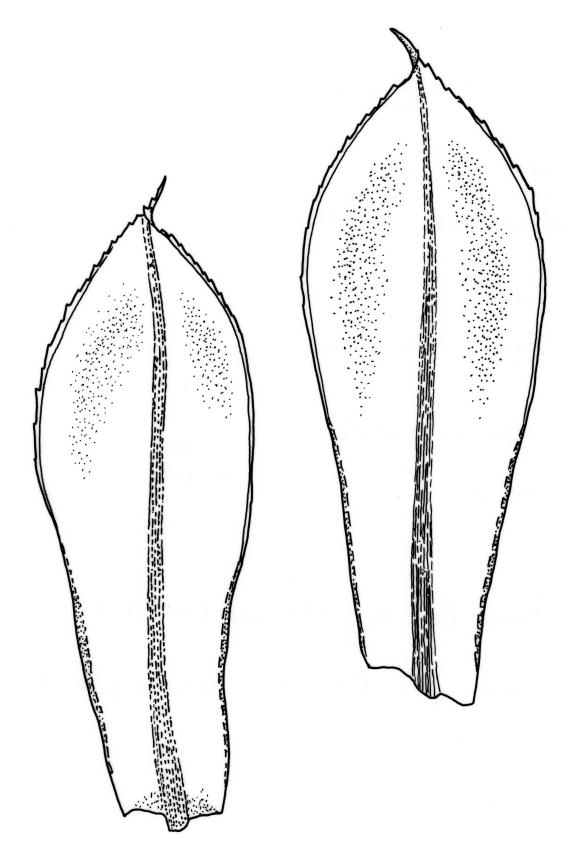
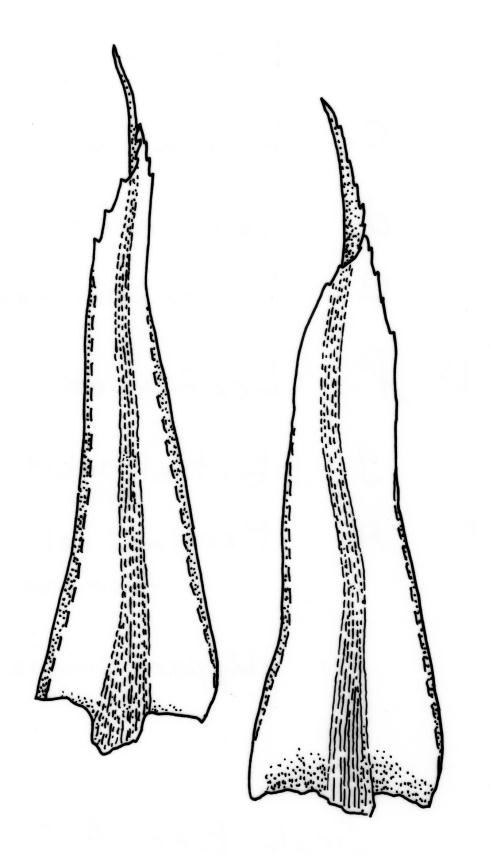
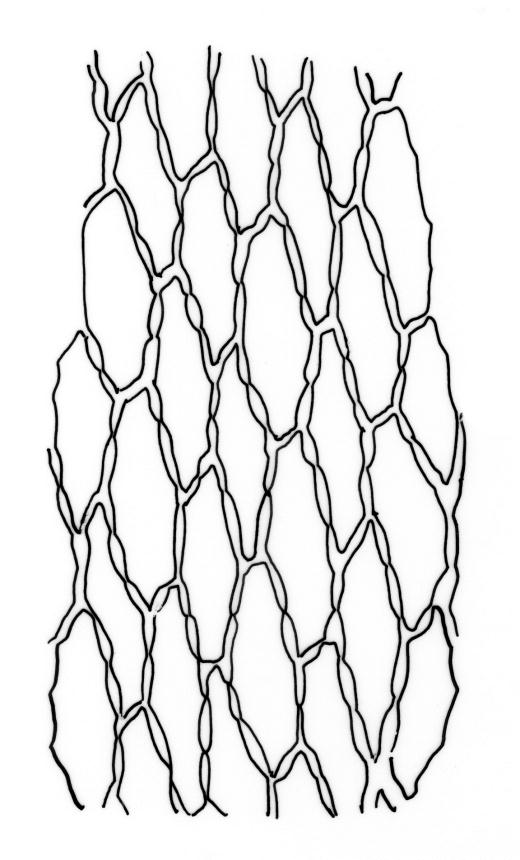
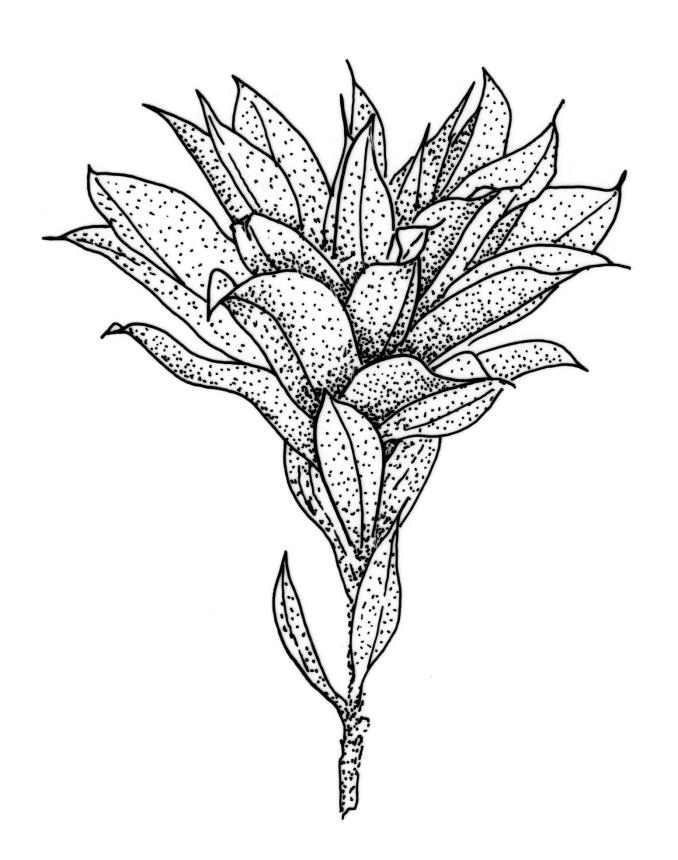
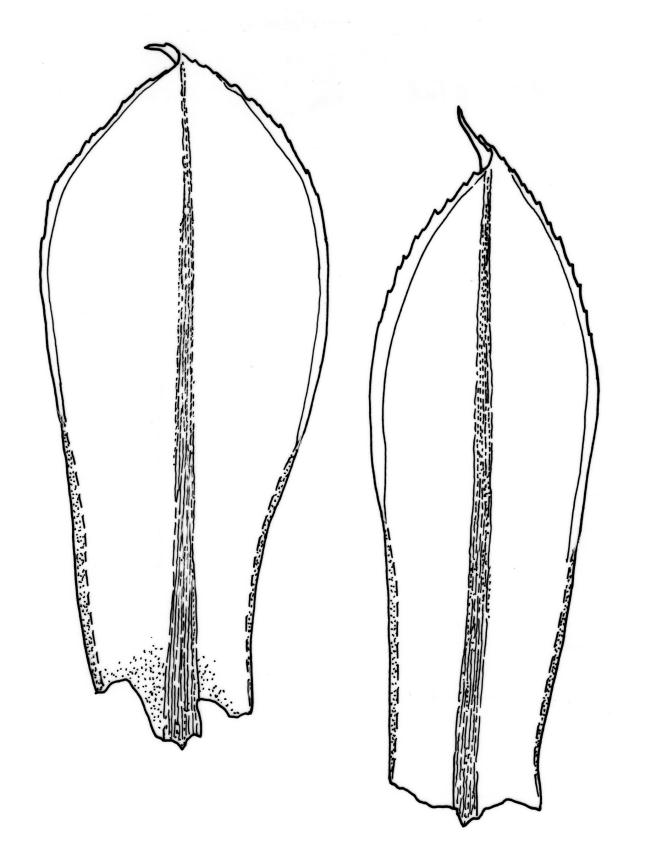
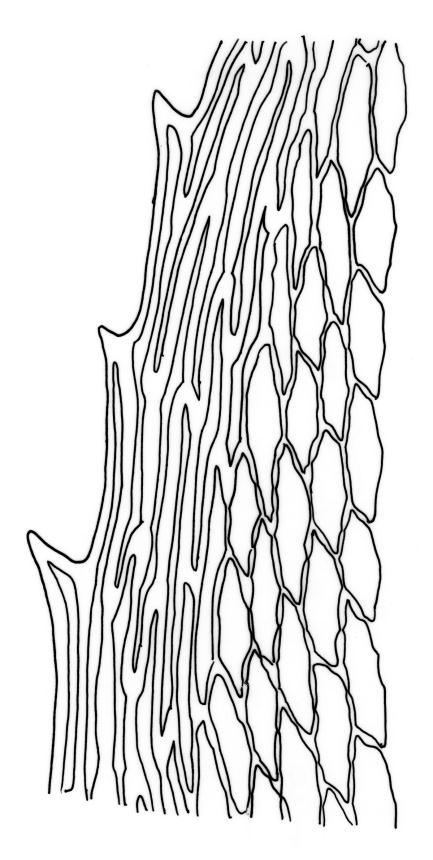
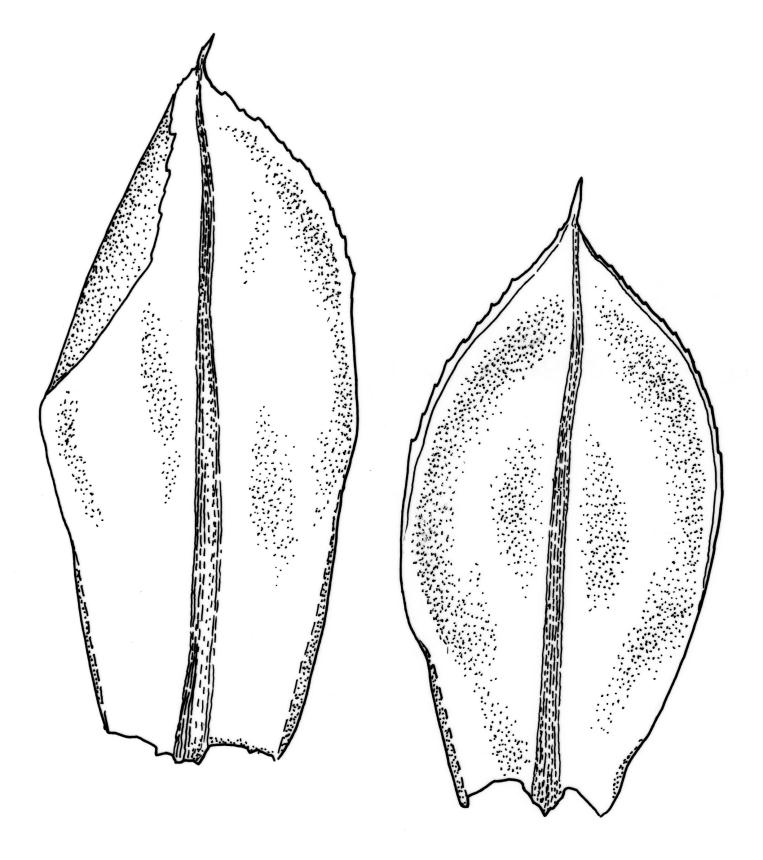
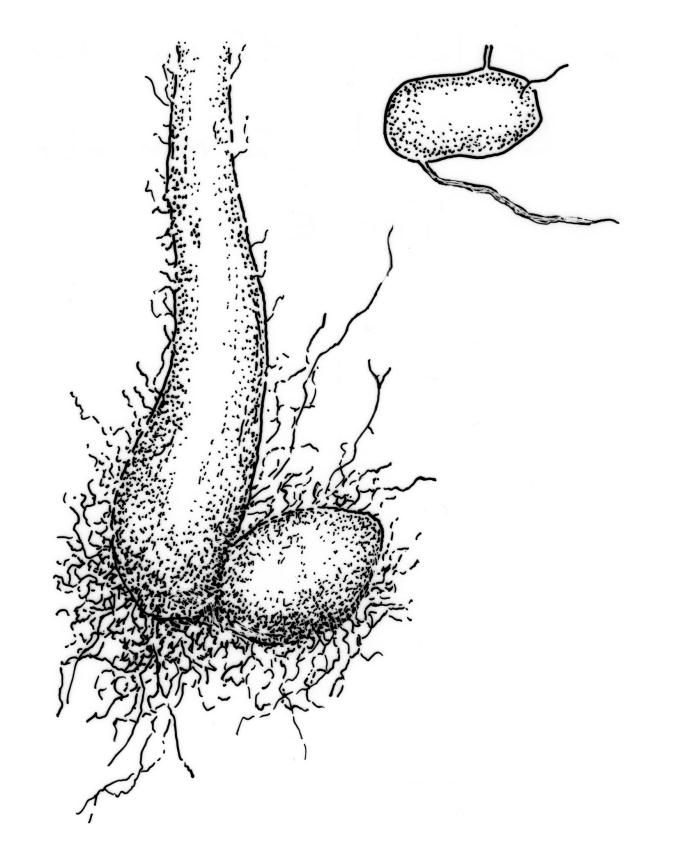
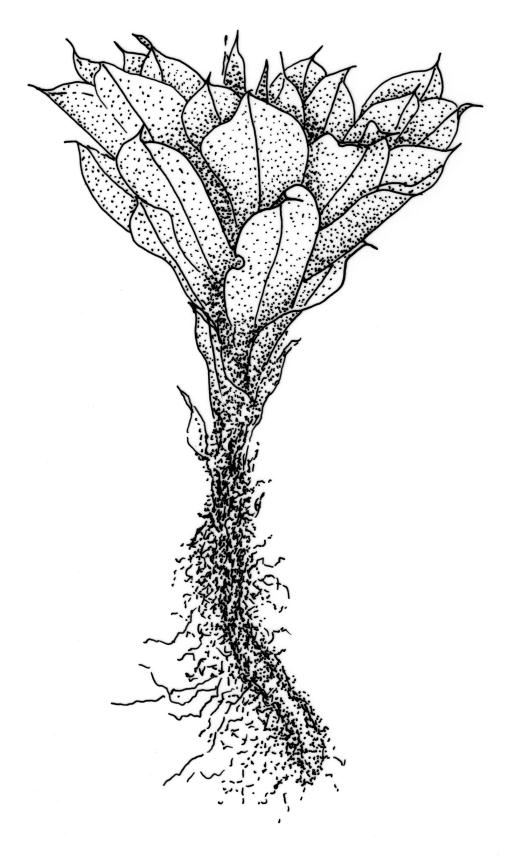
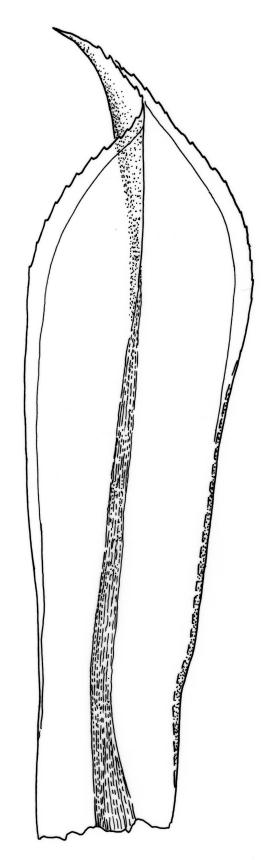
Plants large to robust, bright or yellow-green, weakly lustrous, variable in stature, comose, forming turves. Stems red-brown, c. 10 to at least 35 mm, with innovations arising from within the perichaetia, in cross-section as per genus. Leaves distinctly enlarged and more crowded at stem apices, often serially comose, wide-spreading when moist, much altered, more erect, and often twisted around the leaf axis when dry, obovate to spathulate, widest well above mid leaf, abruptly tapered to an acute, stoutly triangular-cuspidate and variably reflexed apex, c. 3–7 × 1.2–1.5(–1.7) mm (including cusp), sharply serrate or rarely denticulate in upper ⅓ or more, strongly bordered, recurved below, not decurrent; upper laminal cells hexagonal to ± rhombic, firm-walled and porose (often strongly so), c. 45–75 µm long and 3–4:1, in lower leaf becoming longer (to c. 100 µm), more oblong, and usually more porose; marginal cells linear, thicker-walled, and paler to form a wide (commonly 3–8 cells at widest part of leaf) but often poorly defined border, extending to apex and fusing with cells of the cusp; basal cells usually darker brown than adjacent cells. Costa pale brown throughout, excurrent into the triangular, sharply pointed, and ± reflexed cusp. Tubers often present, c. 500–900 µm in greater diam., orange-brown, oval, round, or irregular in outline.
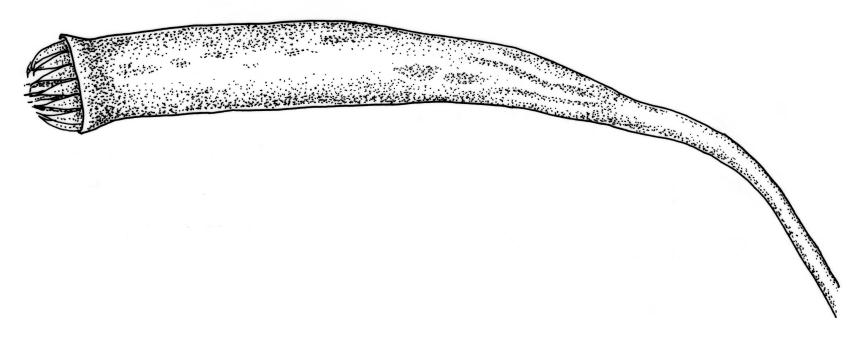
Dioicous. Perichaetia terminal, the inner leaves much smaller than those of the coma, lanceolate or oblong-lanceolate, slenderly acuminate, with a long-excurrent costa. Perigonia terminal and scarcely distinguishable from perichaetia, with broadly ovate-lanceolate inner bracts and red-brown filiform paraphyses. Setae usually multiple (2–4 per perichaetium, less often 1), c. 20–50 mm, red, usually strongly hooked just below capsule; capsules cernuous or pendent, narrowly cylindric, curved, 3–6 mm, with a narrow, scarcely wrinkled, and ± curved neck c. ¼ the capsule length; operculum mammillate, scarcely apiculate. Exostome teeth yellow-brown; endostome with perforate segments nearly the length of the teeth and cilia in groups of 3, mostly appendiculate, but sometimes (variable in single capsule) reduced and nodose. Spores variable in single capsule, 11–18 µm diam., finely papillose.
Mohamed 1979, fig 2, b; fig. 3, b; fig. 5 a–h (as Bryum billardieri var. platyloma); Spence & Ramsay 2006, fig. 49 i–q.
The distinction between R. subtomentosum and R. capillare can be difficult, particularly when lax growth forms of the former are involved. When dry, the leaves of R. subtomentosum tend to spiral around their own axes, while those of R. capillare spiral around the stem. The leaves of R. subtomentosum are mostly sharply serrate rather than entire or denticulate as in R. capillare. Additionally, in R. subtomentosum the leaf border tends to be wider, upper laminal cells more thick-walled and porose, and the apical cusp shorter and stouter than the corresponding structures in R. capillare. Sexuality also distinguishes these taxa in N.Z., as does rhizoidal tuber morphology, when these are present.
Rosulabryum subtomentosum is best distinguished from R. perlimbatum by having narrower leaf borders, stronger and sharper marginal teeth, and larger tubers. Rosulabryum subtomentosum is best distinguished from R. campylothecium by having more spreading leaves with wider borders and reflexed apices, smaller spores, and appendiculate cilia.
NI: N Auckland including offshore islands (TK, PK, HC, LB, GB, RT), S Auckland, Gisborne, Hawke's Bay, Taranaki, Wellington; SI: Nelson, Marlborough, Canterbury, Westland, Otago, Southland; St; Ch; A; C; M.
Probably Australasian. Tasmania*, mainland Australia*. The wider distribution of R. subtomentosum cannot be recorded with confidence. Spence & Ramsay (2006) recorded this taxon from all states of Australia (excluding the Northern Territory). Ochi’s (1970) concept of Bryum billardierei apparently corresponds to R. subtomentosum as interpreted here and he recorded it also from New Caledonia and Fiji.
On duff, humus, or mineral soil (especially sand), rotten logs, and shaded rock in forest or scrub, including both coastal and inland situations. R. subtomentosum is well-developed in a wide range of forest types. In the far north it occurs commonly under Leptospermum/Kunzea and Metrosideros forest, often in coastal situations, as well as mixed podocarp-broadleaved forest (as in the Waitakere Ranges). In the central North I. and South I. it is widespread in montane southern beech (especially Fuscospora cliffortioides and F. solandri) forest. It also occurs in exotic (e.g. eucalypt and pine) plantations and rarely in grasslands (and then usually appearing lax in habit). It does not normally extend above tree line. Occurring from sea level to c. 1200 m elevation on both North I. and South I.
When well-developed, R. subtomentosum is easily recognised. Among the most constant features that facilitate its recognition are the hooked setae, the elongate, slender, curved, and usually multiple capsules, and the ± reflexed nature of the leaf apices. The very large tubers, while not always present, are also distinctive. The usually markedly comose habit, especially in plants from drier and more insolated sites, is correlated with the above features, as is the weakly defined, but wide leaf border. Plants growing in very moist habitats sometimes have leaves more evenly spaced along the stem. The features noted in the key will allow placement of nearly all fruiting specimens.
Many of the features of this species, including the degree of differentiation of the comal leaves, the degree of reflexion of the leaf apices, and the breadth of the leaf border, seem to vary independently and a moderate number of collections, particularly sterile ones, cannot be placed confidently as either R. subtomentosum or R. billardierei.
The range of intermediate material involves many permutations of character states, including plants with gametophytes characteristic of R. subtomentosum that bear sporophytes characteristic of R. billardierei (single relatively short, pendent, and nearly symmetric capsules), as in K.W. Allison 919 ex Flagstaff Hill, Otago (CHR 548195).
Mohamed’s (1979, p. 414) suggestion that a progressive reduction in size, border width, and acuminate nature of [the leaf] apex occurs as one proceeds northwards (i.e., from Southland L.D. northwards) in R. subtomentosum (cited as Bryum billardieri var. platyloma) cannot be confirmed. However, plants from N Auckland L.D. generally have more weakly developed comae and leaves less spreading than those from other regions. Also, shorter and broader leaves are correlated with narrower leaf borders throughout both main islands.
Despite the difficulty associated with distinguishing R. subtomentosum and R. billardierei, the recognition of both taxa here is a pragmatic solution to an intractable taxonomic problem. Study using molecular techniques might yield an improved understanding of the population variability in Rosulabryum. Continued study of morphology alone is unlikely to improve our understanding.
The taxonomic confusion surrounding R. subtomentosum and its allies has been greatly compounded by nomenclatural confusion. In particular, the confusion has centred on the names Bryum platyloma Schwägr., Bryum billardierei Schwägr., and, much more recently, Bryum billardierei var. platyloma Mohamed. Bryum platyloma Schwägr. is based on a specimen from Madeira with an unspecified collector. Bryum billardierei Schwägr. is based on a de Labillardière collection of unknown (but probably Mediterranean) provenance; it is discussed elsewhere in this work. Confusingly, the names Bryum platyloma Schwägr. and Bryum billardierei Schwägr. were published on consecutive pages of Sp. Musc. Frond. Suppl. and both taxa were illustrated in the same plate. The name Bryum platyloma Schwägr. was not, to my knowledge, applied in print to Australasian material prior to its inclusion in the synonymy of R. subtomentosum (Hampe) J.R. Spence by Spence & Ramsay (2006). Their statement (p. 344) that Bryum platyloma Schwägr. and Bryum billardierei var. platyloma Mohamed are homotypic (nomenclatural) synonyms is incorrect.
Much of the reason for this nomenclatural confusion can be traced to Mohamed’s choice of an epithet for a plant he considered to be a variety of Bryum billardierei Schwägr. The name B. billardierei var. platyloma Mohamed has been widely applied to a common and widespread N.Z. (and Australian) taxon since its publication in 1979. Mohamed’s name is validly published. He clearly and correctly specified John Child 2713 as the type of the var. platyloma. The salient features of Mohamed’s type include the length and number of the capsules, the hooked nature of the upper setae, and the capsule form and length. Spence & Ramsay’s (2006) designation of J. Child 2713 as a neotype is superfluous.
In a N.Z. context, recognition of this taxon at the species, rather than variety level, is advocated here. At the species level, the name R. subtomentosum (Hampe) J.R.Spence is preferred since, as determined by Spence & Ramsay (2006), its Victorian basionym is the earliest valid name at species rank. Recognition at species level using this name obviates taxonomic and nomenclatural confusion with R. billardierei (Schwägr.) J.R.Spence. The last taxon has (as discussed elsewhere in this work) particular problems surrounding the provenance of its type.
Rosulabryum subtomentosum resists neat taxonomic treatment; a specialised study using non-morphological characters is probably the only way to improve our understanding of this species and its congeners.