Class
Family
Dicranoloma pungentella sensu Dixon 1913, p. 26. See discussion below.
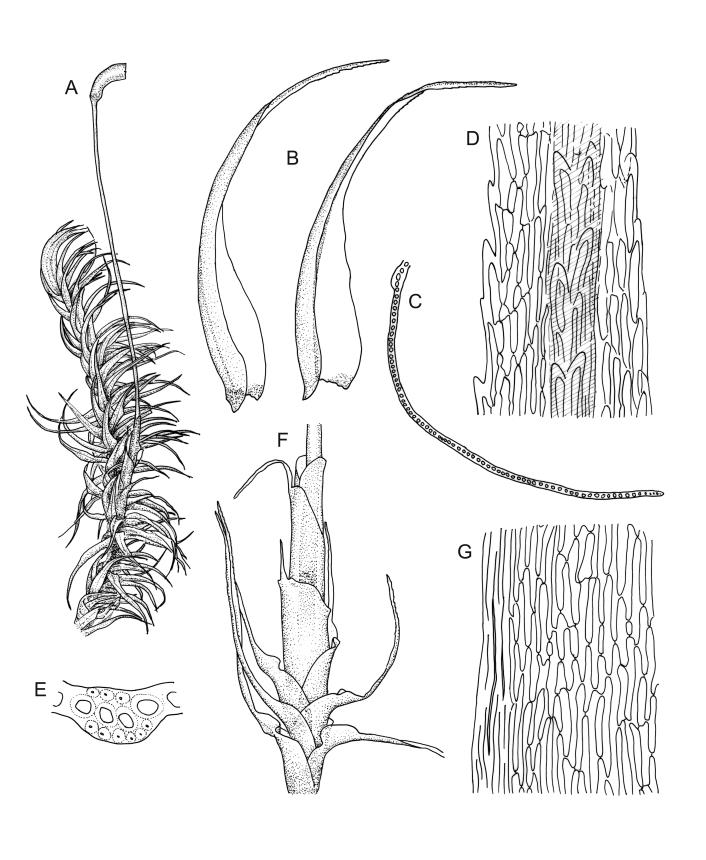
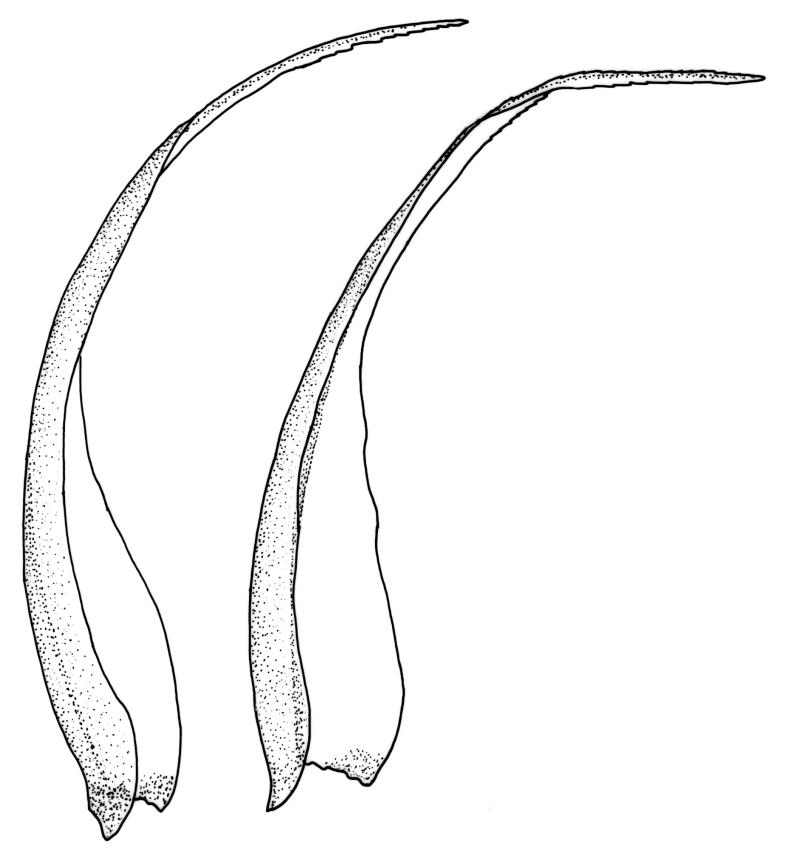
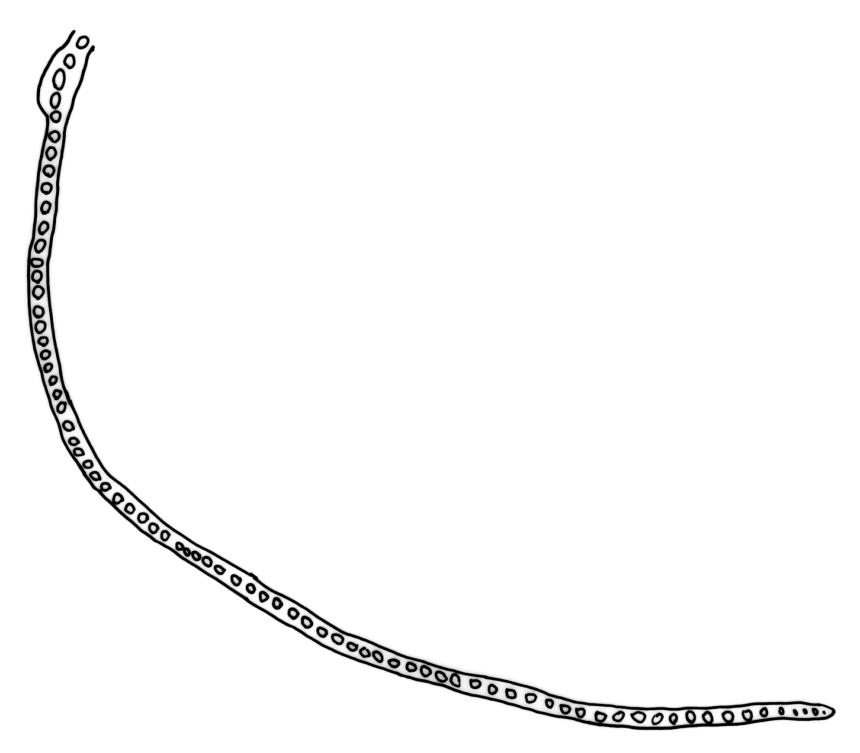
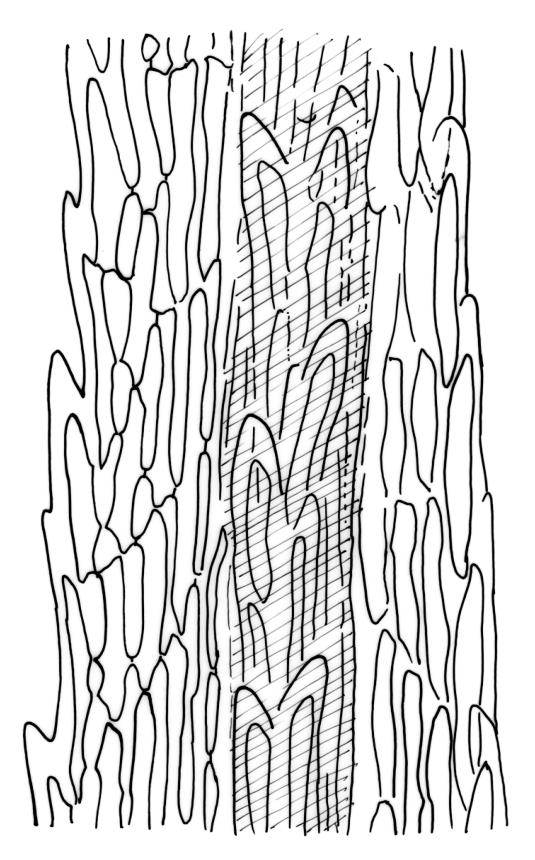
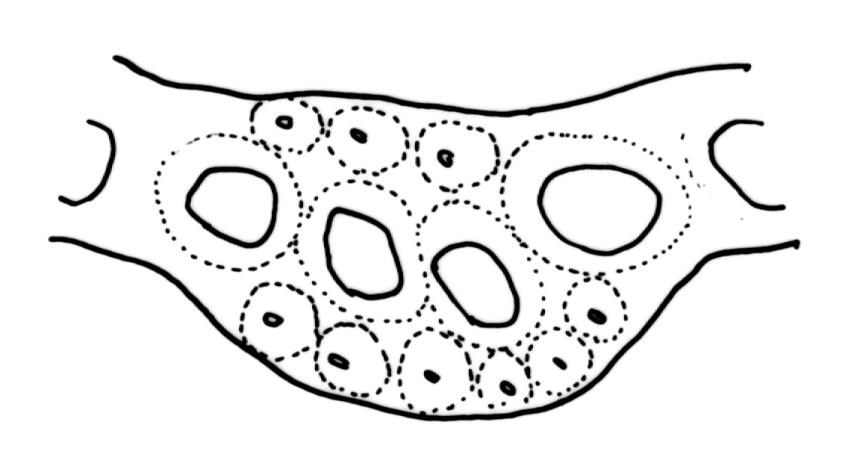
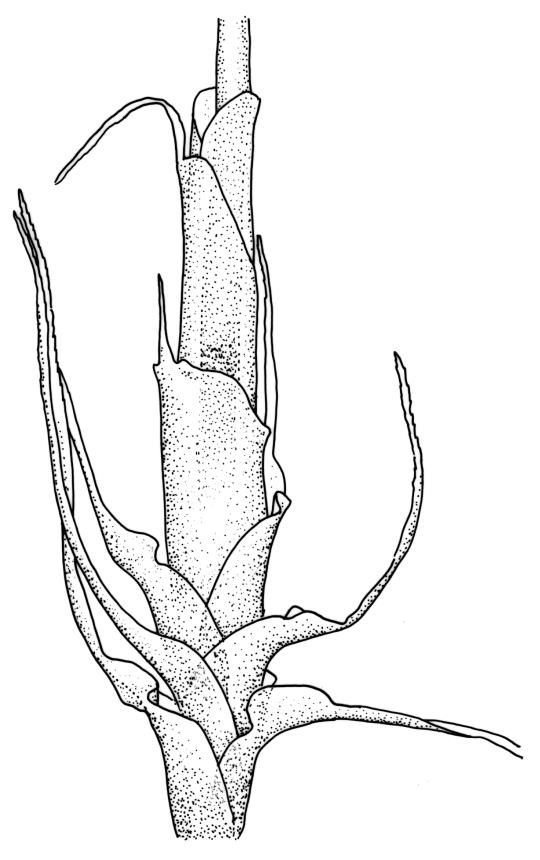
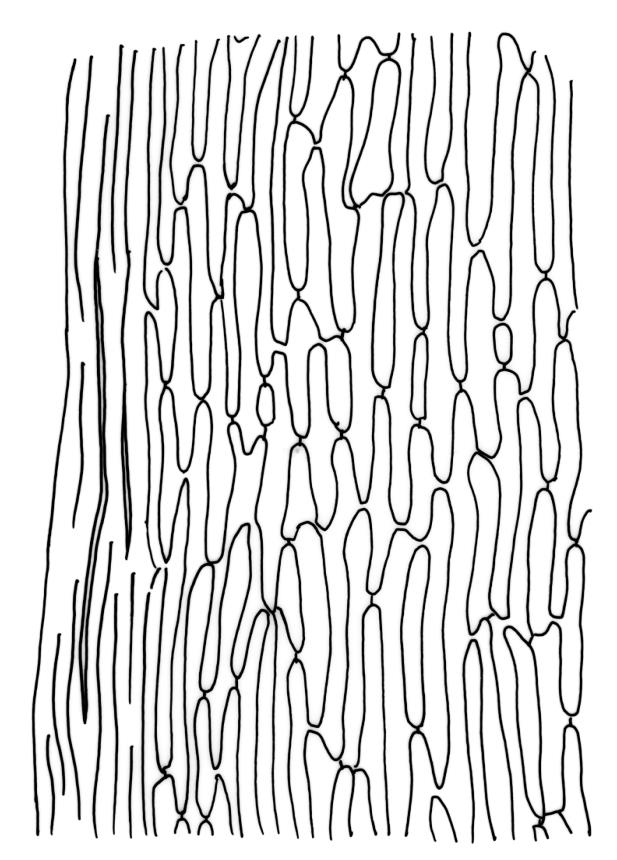
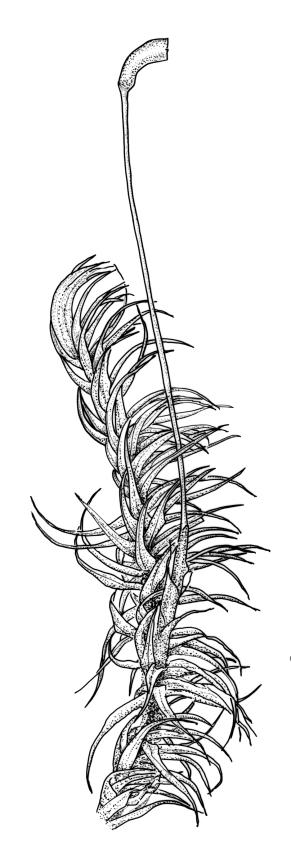
Plants medium-sized, pale yellow-green and lustrous when fresh, becoming gold-brown in older dried specimens, mostly forming cushions. Stems very conspicuous in fresh material, 30–100 mm, branched by both forking and innovation, in cross-section with 3–4 layers of thick-walled cortical cells and an ill-defined central strand, with sparse pale rhizoids in leaf axils. Leaves falcate-secund or erect-spreading when fresh, often twisted at apex but neither plicate nor rugose when dry, ovate-lanceolate, ± narrowed to insertion, serrate to ± entire above (usually serrate in upper ¼ or less, and teeth one-celled), entire below, (4–)5–8 × c. 1.5 mm (under cover slip), subtubulose, plane at margins; mid laminal cells irregular and elongate, mostly 39–75 × 4–6 μm and 8–15:1, porose, with elongate cells extending to apex; juxtacostal cells at mid leaf not differentiated; border well defined and extending from just above alar group to serrations, mostly 3–4 cells and c. 15(–24) µm wide at mid leaf; cells of leaf base slightly longer (to c. 105 µm) but otherwise as those of mid lamina; alar cells extending c. ½ to costal base and c. 10 cells up the margin, brown, not or slightly inflated. Costa narrow, 30–45 µm wide at mid leaf, occupying c. 1/25 or less of widest part of leaf base, the abaxial surface in c. upper ⅕ bearing a single or double row of short spines (not visible under hand-lens), in cross-section lacking wings at mid leaf, with 3–5 guide cells in 1–2 layers, with (4–)6–8 abaxial stereids in a ± single surface layer (sometimes ± divided into 2 groups by a guide cell) and 2–5 stereids on the adaxial surface.
Pseudautoicous. Perichaetial leaves sheathing (c. ¼ the seta length), ecostate, obtuse or retuse, mucronate (mucro sometimes to c. 0.1 the total leaf length or rarely longer). Setae one per perichaetium, 19–30 mm, red-brown; capsules exserted, strongly curved, c. 2.8–3.5 mm, strumose, smooth both when moist or dry, otherwise as per genus; operculum curved-rostrate from a conic base, ± equal to the capsule. Peristome teeth c. 350–440 × 90 µm, otherwise as per genus. Calyptra cucullate, smooth. Spores 17–21 µm.
Dixon 1913, Pl. IV, figs. 12, 14–15, 19 (variously named); Tan & Koponen 1983, figs 41–43; Beever et al. 1992, fig. 24 a–e; Klazenga 2003, figs 3 a–p & 4 a–d; Seppelt 2004, fig. 54.
Dicranoloma billardierei often grows in association with D. robustum. The shorter leaves, generally more branched stems, and obtuse (vs long aristate) perichaetial leaves are all useful to distinguish the two species under field conditions. D. billardierei colonies are nearly always paler and less yellow than those of D. robustum; this is a subtle difference but is obvious in the field. However, D. billardierei lacks the distinct white-green colour of such species as D. dicarpum, D. fasciatum, and D. plurisetum. According to Seppelt (2004), sterile plants of D. billardierei and D. robustum on Macquarie I. can be separated by the number of guide cells observed in a costal cross-section (presumably mid leaf): two to four in the former and three to eight in the latter; this largely accords with my observations, at least as far as "representative" material is concerned.
Some of the confusion between D. billardierei and D. robustum can be traced to Scott & Stone’s (1976, p. 154) opinion that the two species were not separable.
In addition to the aforementioned features, mid leaf costal cross-sections can be used to distinguish the two species. In D. billardierei the costa at mid leaf has 3–5 guide cells, 6–8 abaxial stereids aligned in a single layer (which extends to the surface), 5 or fewer adaxial stereids, and seldom if ever exceeds 45 µm in width. There are no cells with larger lumens (relative to the stereids) on the abaxial surface. The costal structure corresponds to Renauld’s heteroneuron (occasionally leptoneuron) costal type. In D. robustum the costa at mid leaf has 6–12 median guide cells with mostly two layers of stereids abaxially and adaxially, and 60–160(–225) µm wide. Sometimes the surface cells on the abaxial side are larger lumened than the adjacent stereids. The costal structure of D. robustum corresponds to Renauld’s toxoneuron costal type. The costal structure of D. billardierei is more uniform than that of D. robustum.
In unpublished work done in 1988, R. Lewington (pers. comm.) found that the average length:width ratio of fully developed D. billardierei vegetative leaves ranged from 3.2–7.6:1 in contrast to a ratio of 5–13.2:1 for D. robustum. Lewington measured six representative leaves per herbarium specimen, and used predominantly herbarium specimens named by Sainsbury.
When sterile, D. billardierei is sometimes confused with D. fasciatum. Dicranoloma billardierei has serrate rather than spinose-serrate upper leaf margins. The marginal toothing and spines on the abaxial surface of the costa in D. billardierei are scarcely visible under the hand-lens. The leaf border in D. billardierei is c. 15–24 µm wide, while that of D. fasciatum is wider (30–75 µm) and clearly visible under the hand-lens. The lack of leaf plications in D. billardierei also serves to distinguish it. When fruiting the two species are unlikely to be confused.
NI: N Auckland, including offshore islands (PK, LB, GB, RT), S Auckland, Gisborne, Hawke’s Bay, Taranaki, Wellington; SI: Nelson, Marlborough, Canterbury, Westland, Otago, Southland; St; Ch; A, C. Reported from M by Seppelt (2004). Relatively few collections have been confirmed from Hawke’s Bay and Otago L.D.
Austral. Tasmania*, mainland Australia*, Marion I, South Africa*, Tanzania*, Argentina*. Recorded from Madagascar and southern South America by Klazenga (2003) and from Juan Fernandez Is. Robinson (1975).
Both terrestrial and epiphytic. Growing on soil or rock hummocks, logs, stumps, and exposed roots; less often directly on rock. Terrestrial in a variety of scrub and forest types and often on duff in well-drained sites. Occurring on the forest floor of southern beech or mixed broad-leaved forest, and in N Auckland often occurring in mānuka scrub. Very often forming a mosaic with D. robustum. From near sea level (Rangitoto I.) to c. 1140 m (north-western Ruahine Range, Wellington L.D.) on the North I. and from near sea level (Pororarī River, Westland L.D.) to at least 1250 m (Temple Basin, Canterbury L.D.) on the South I.
The compact cushions and the combination of stems, which are clearly visible among the leaves and much-branched, the tendency for some stems to become prostrate, and the relatively short leaves make this species distinctive, particularly in the field. The leaves are usually falcate-secund, but forms with erect-spreading leaves are not rare, particularly in boggy habitats. The conspicuousness of the stems is largely due to the much sparser production of axillary rhizoids in this species than in most of its N.Z. congeners. The absence of plications in the lower leaf helps to distinguish it from several species. Capsules, when present, are invariably one per perichaetium. The obtuse (and sometimes mucronate) nature of the innermost perichaetial leaves is distinctive and readily observed under a hand-lens. When growing terrestrially D. billardierei usually forms pure cushions of up to c. 0.8 m or more in diameter.
Dicranum billarderi var. duriusculum Hook.f. & Wilson was reduced to synonymy by Klazenga (2003). Dicranum orthopyxis Müll.Hal. and Dicranum scopelloides Paris were placed in synonymy by Dixon (1913, p. 23) and tentatively accepted by Klazenga (2003). These synonymies are followed here without further investigation.
The name D. angustinerve (Mitt.) Paris [Index Bryol., ed. 2, 2: 24, 1904)] has a Tasmanian type. It was applied by Mitten (in herb. Beckett) to material from Great Barrier I. Mitten’s (1859) protologue described the species (as a Dicranum) as having inner perichaetial leaves "furnished with a bristle-like point, which seems wanting in D. billardierei." The Great Barrier I. material (T. Kirk 46, CHR 543127) indeed has strongly aristate inner perichaetial leaves but otherwise falls within the range of continuous variation for D. billardierei and in my opinion is referable here. Klazenga (2003) studied Tasmanian type material of Mitten’s name and unequivocally placed it in the synonymy here.
The Tasmanian name D. pungentella (Müll.Hal.) Paris has been applied to N.Z. material by Dixon (1913, p. 26), Sainsbury (in herb.) and others. N.Z. material so-named has leaves more spreading and less secund and upper margins somewhat less serrate than usual for D. billardierei. The vegetative leaf length is usually near the lower end of the range of continuous variation for this species. In at least some instances these minor variations are associated with occurrences in boggy sites; they are not significant in terms of the overall variability of D. billardierei. The N.Z. material named as D. pungentella by Sainsbury (no material named as D. pungentella by Dixon has been seen) is from epiphytic habitats and sterile, and has weakly secund and weakly toothed leaves of c. 5–6 mm. This material is quite unremarkable in the context of N.Z. D. billardierei. Klazenga (2003) tentatively accepted Sainsbury’s (1955) treatment of D. pungentella (Müll.Hal.) Paris as a synonym of D. billardierei but he was unable to locate type material from Mt Wellington, Tasmania.
Dicranoloma integerrimum (Broth. & Geh.) Paris was likewise recorded from a single Stewart I. locality by Dixon (1913) and synonymised with D. billardierei by Sainsbury (1955). However, it has been more convincingly synonymised with D. robustum by Klazenga (2003), who located and examined Tasmanian type material. Klazenga’s placement of this name is accepted here. The Stewart I. collection (leg. Cockayne) has not been located.
Dixon (1913, p. 23) ascribed additional names, based on Australian types (which have not been applied to N.Z. material), to the synonymy of D. billardierei; some of these synonyms have been typified and confirmed by Klazenga (2003). Tan & Koponen (1983) provided additional synonymy from across a wide geographic range, including names from Malesia and South America; Klazenga (1999) agreed with Tan & Koponen in regard to Malesian and south-east Asian synonymy. Some additional synonymy was provided by Robinson (1975, p. 22), but it is unclear how many of the types he cited were examined by him.
Material of Dicranum pelliceum Müll.Hal. nom. nud. (H-Br 1201039!) and D. turgidum Müll.Hal., nom. nud. (CHR 543125!) is referable to Dicranoloma billardierei.