
- Taxon
- Gallery
Classification
Nomenclature
Scientific Name:
Veronica lilliputiana Stearn, Gard. Chron., ser. 3, 129: 166, No. 3359 (1951)
Synonymy:
- ≡ Veronica canescens Kirk, Trans. & Proc. New Zealand Inst. 9: 503, t. 19 (1877) nom. illeg., non Veronica canescens Schrad. 1803
- ≡ Hebe canescens A.Wall, Trans. & Proc. New Zealand Inst. 60: 384 (1929) nom. nov. pro Veronica canescens Kirk 1877
- ≡ Parahebe canescens (A.Wall) W.R.B.Oliv., Rec. Domin. Mus. 1: 229 (1944)
Holotype: T. Kirk, Lake Lyndon, WELT 5112a&b
Etymology:
Description
Short-lived, perennial herb, older stems sometimes slightly woody, to 0.01 m tall. Stems creeping, with short, sub-erect axillary shoots, eglandular-pubescent or rarely glabrous; hairs uniform. Leaf bud indistinct; leaves separating while very small, sub-distichous on prostrate shoots, spreading to recurved, thin, ovate, elliptic, obovate, or orbicular, 1–3 mm long, 1–2 mm wide, dull and glaucous above and beneath, veins not evident, surfaces eglandular-hairy; margins ciliate, entire; apex obtuse to rounded; base cuneate; petiole 0–0.5 mm long. Flowers solitary or rarely paired, on short, bibracteate peduncles in axils of opposite leaves, all bisexual; bracts opposite, elliptic to ovate, < pedicels; pedicels erect, 2–7 mm long, eglandular-hairy all around. Calyx lobes 4, rarely 5, sub-acute to obtuse, equal, 2–3 mm long, eglandular-hairy. Corolla c. 10 mm diameter; tube white and yellow, c. 1 mm long, < calyx, eglandular-hairy inside; lobes 4, rarely 5 by division of posterior lobe, pale to purplish-blue, sub-erect, sub-equal, elliptic to narrow-elliptic, 3–6 mm long, obtuse; nectar guides dark blue. Stamen filaments white, 4–5 mm long; anthers purplish. Style glabrous, 4.0–4.5 mm long. Capsules angustiseptate, emarginate, 1.0–1.8 mm long, 1–2 mm at widest point. Seeds ellipsoid to obovoid, weakly flattened, finely reticulate, pale brown to brown, 0.4–0.6 mm long.
Recognition
V. lilliputiana is unlike any other New Zealand species of Veronica. Plants distinctively have a combination of creeping habit, tiny, entire glaucous and hairy leaves, and blue, usually solitary, bibracteate flowers.
Tetrachondra hamiltonii plants (Lamiaceae) have a similar size and growth form, but leaves are brighter green, glabrous or very minutely ciliolate, and gland-dotted; calyx and corolla five-lobed; corolla two-lipped; fruit four nutlets.
Distribution
South Island: Canterbury, Otago, Fiordland (Lake Manapouri only). Mostly inland, but known also from a few coastal sites.
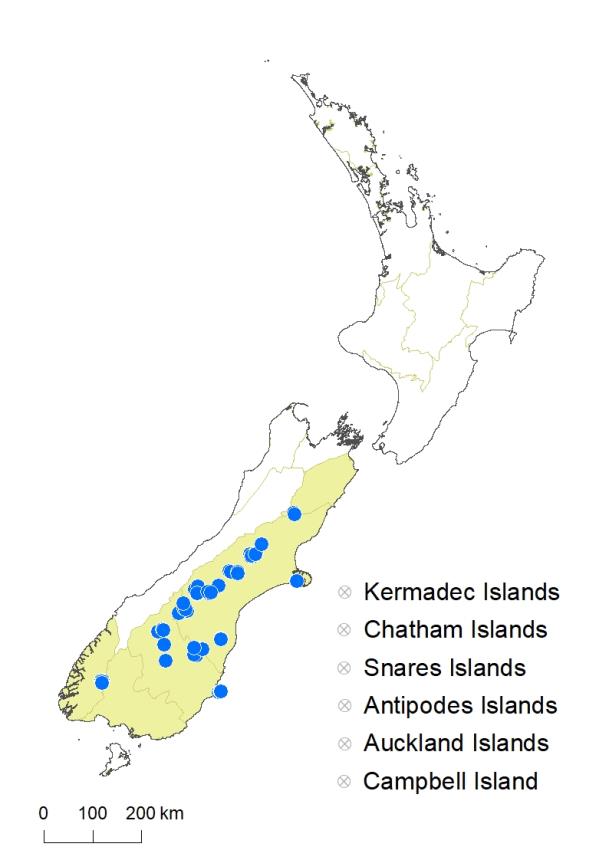
Habitat
Seasonally wet (usually summer dry) margins of coastal to montane lakes, tarns, kettle-holes, ponds, and depressions in moraines, light soil among stones or in turf of other small plants such as Leptinella maniototo and Galium perpusillum. Recorded elevations range from 20 to 1441 m.
Biostatus
Indigenous (Endemic)
Phenology
Flowers: December–April; fruits: January–May.
Cytology
2n = 42 (Hair 1970, as Parahebe canescens).
Notes
Veronica lilliputiana is classified in V. subg. Pseudoveronica sect. Hebe (Albach & Meudt 2010). It appears to belong with V. lyallii and several other species in a clade (the speedwell hebes, Albach & Meudt 2010) that is characterised by short corolla tubes, nectar guides on the corolla, and rather lax inflorescences. Most also have plicate lateral corolla lobes, which enfold the stamens, but V. lilliputiana lacks these, along with V. linifolia, V. colostylis, and V. jovellanoides.
In V. lilliputiana, vegetative reproduction is possible because small, short branches detach easily and may be transported in water to other sites, where they can establish by adventitious roots. These reduced branches have 2(–3) pairs of close-set leaves. They may make it easy for V. lilliputiana to spread rapidly around the margins of lakes and tarns.
Images




Bibliography
Albach, D.C.; Meudt, H.M. 2010: Phylogeny of Veronica in the Southern and Northern Hemispheres based on plastid, nuclear ribosomal and nuclear low-copy DNA. Molecular Phylogenetics and Evolution 54: 457–471.
de Lange, P.J.; Gosden, J.; Courtney, S.P.; Fergus, A.J.; Barkla, J.W.; Beadel, S.M.; Champion, P.D., Hindmarsh-Walls, R.; Makan, T.; Michel, P. 2024: Conservation status of vascular plants in Aotearoa New Zealand, 2023. New Zealand Threat Classification Series. No. 43. [Declining]
de Lange, P.J.; Rolfe, J.R.; Barkla J.W.; Courtney, S.P.; Champion, P.D.; Perrie, L.R.; Beadel, S.N.; Ford, K.A.; Breitwieser, I.; Schönberger, I.; Hindmarsh-Walls, R.; Heenan, P.B.; Ladley, K. 2018: Conservation status of New Zealand indigenous vascular plants, 2017. New Zealand Threat Classification Series. No. 22. [Declining]
de Lange, P.J.; Rolfe, J.R.; Champion, P.D.; Courtney, S.P.; Heenan, P.B.; Barkla, J.W.; Cameron, E.K.; Norton, D.A.; Hitchmough, R.A. 2013: Conservation status of New Zealand indigenous vascular plants, 2012. New Zealand Threat Classification Series 3. Department of Conservation, Wellington. [as Parahebe canescens (A.Wall) W.R.B.Oliv.] [Declining]
Garnock-Jones, P.J. 2023: Veronica. In: Breitwieser, I. (ed.) Flora of New Zealand – Seed Plants. Fascicle 9. Manaaki Whenua Press, Lincoln.
Garnock-Jones, P.J.; Albach, D.; Briggs, B.G. 2007: Botanical names in Southern Hemisphere Veronica (Plantaginaceae): sect. Detzneria, sect. Hebe, and sect. Labiatoides. Taxon 56: 571–582.
Garnock-Jones, P.J.; Lloyd, D.G. 2004: A taxonomic revision of Parahebe (Plantaginaceae) in New Zealand. New Zealand Journal of Botany 42: 181–232.
Hair, J.B. 1970: Contributions to a chromosome atlas of the New Zealand flora — 13. Parahebe and Pygmea (Scrophulariaceae). New Zealand Journal of Botany 8: 255–259.
Kirk, T. 1877: Description of two new species of Veronica. Transactions and Proceedings of the New Zealand Institute 9: 502–503.
Oliver, W.R.B. 1944: The Veronica-like species of New Zealand. Records of the Dominion Museum, Wellington 1: 228–231.
Stearn, W.T. 1951: Veronica lilliputiana. Gardeners' Chronicle, ser. 3, 129: 166.
Wall, A. 1929: A preliminary catalogue of New Zealand plants cultivated in Britain. Transactions and Proceedings of the New Zealand Institute 60: 379–393.