Class
Subclass
Order
Family
Genus
Rhizomes long-creeping, 1–2 mm diameter, with stipes arising 15–90 mm apart; bearing red-brown non-glandular hairs up to 3.5 mm long. Fronds 240–1100 mm or rarely to 1900 mm long. Stipes 85–700 or rarely to 900 mm long, 1.5–5 mm diameter, dark purple-brown to red-brown, bearing abundant red-brown non-glandular hairs up to 3 mm long proximally, becoming scattered distally and replaced by abundant colourless glandular hairs, which are variable in length but shorter than the brown hairs. Rachises red-brown proximally, becoming yellow-brown distally, densely covered in short colourless glandular hairs 0.1–0.8 mm long interspersed with scattered red-brown non-glandular hairs to 2 mm long. Laminae 2-pinnate-pinnatifid to 3-pinnate-pinnatifid, ovate, tapering to a long pinnatifid apex, 145–1000 mm long, 115–600 mm wide, light green on both surfaces, herbaceous, exuding white milky substance from adaxial surface with age; lamina margins and abaxial lamina surfaces and costae densely covered in colourless glandular hairs, 0.1–0.3 mm long on laminae, up to 0.4 mm long on costae, interspersed with a few colourless non-glandular hairs of similar length; occasional red-brown non-glandular hairs up to 1 mm long on costae. Primary pinnae in 20–35 pairs below long-tapering pinnatifid apex, widely spaced or slightly overlapping, the proximal pair arising at 35–80° to rachis, winged distally; distal primary pinnae narrowly ovate to narrowly oblong; proximal primary pinnae ovate to triangular; the longest at or near the base, 65–385 mm long, 32–155 mm wide, apices long-acuminate, bases short-stalked. Secondary pinnae gradually decreasing in length along each primary pinna to the distal end, winged throughout; the longest narrowly ovate or narrowly oblong, 17–90 mm long, 8–30 mm wide, apices acute or obtuse, bases short-stalked. Tertiary pinnae winged throughout; the longest oblong, 4–16 mm long, 2–7 mm wide, apices obtuse, bases adnate; margins deeply divided on larger fronds into ultimate segments up to 4 mm long and 2.5 mm wide. Veins ending in apices of ultimate segments, sometimes slightly excurrent. Sori ± round, virtually unprotected; paraphyses absent. Mean spore size 28–35 μm long, 18–21 μm wide; perispores pale, echinate and reticulate.
Hypolepis lactea is one of three similar species in New Zealand that have glandular hairs, red- or purple-brown stipes and rachises, and lack indusial flaps. It is distinguished from H. rufobarbata by the presence of colourless glandular and non-glandular hairs on the lamina margin in contrast to the abundant red-brown acicular hairs in H. rufobarbata. Hypolepis lactea is further distinguished by its milky exudate on the adaxial lamina surface at maturity, and by its veins, which are sometimes slightly excurrent. It also has abundant short, colourless glandular hairs on the rachises and costae, in contrast to H. rufobarbata, which has red-brown glandular or acicular hairs. Hypolepis lactea is more similar to H. amaurorhachis but is distinguished by its darker purple-brown stipe and rachis, which become green only at the far distal end, and by its shorter glandular hairs on the laminae (0.1–0.3 cf. 0.3–0.7 mm long). The distributions of the two species do not overlap, H. lactea being a plant of the North Island and South Island except for the far south, whereas H. amaurorhachis occurs only in the far south of the South Island and on the subantarctic islands.
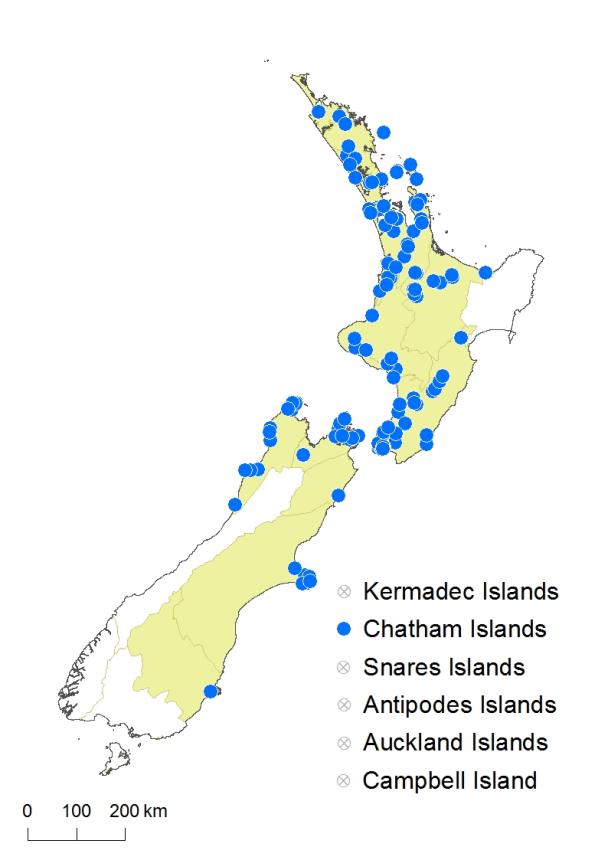
North Island: Northland, Auckland, Volcanic Plateau, Taranaki, Southern North Island.
South Island: Western Nelson, Sounds-Nelson, Marlborough, Canterbury, Otago.
Chatham Islands.
Altitudinal range: 0–800 m.
In the North Island Hypolepis lactea occurs from near Kaitāia to Wellington but is found most frequently in coastal and lowland sites from Kaipara Harbour southwards, extending locally into montane areas and reaching 800 m near Rotorua. It is absent from the central high country, Gisborne Ecological Province and much of the east coast. In the South Island it occurs in coastal and lowland areas of the Marlborough Sounds and north-west Nelson as far south as Greymouth, reaching 600 m near Denniston. On the east coast there are isolated populations near Kaikōura, on Banks Peninsula and around Dunedin. It is common on the Chatham Islands.
Grows on swampy and peaty soils, on the bases of Carex secta in bogs, on disturbed or stony ground in forest and scrub, in clearings, on bush margins, clay banks, stream banks, drainage ditches, and roadsides, at the base of uprooted trees, and on rotting logs. It occurs in kauri, podocarp, broadleaved and beech forest, and under mānuka. It is similar in its ecological tolerances to H. rufobarbata, but occurs generally at lower altitudes.
There is evidence for hybridisation between H. lactea and H. ambigua (AK 298766–298767) and H. dicksonioides (WELT P011537). Hybrids can be recognised by their aborted spores (see Brownsey & Chinnock 1984). There is also some morphological evidence that H. lactea and H. rufobarbata hybridise when they occur together (AK 270457–60, CHR 323893, WELT P011467), but no aborted spores have been found and further work is needed to determine whether such hybrids exist and whether they retain some degree of fertility (Brownsey & Chinnock 1984).
n = 52 (Brownsey & Chinnock 1984).
In New Zealand this species has been widely misidentified by earlier authors as Polypodium rugosulum Labill., P. punctatum Thunb., or combinations based on them.
Hypolepis lactea is morphologically and cytologically distinct from the Australian H. rugosula, which has n = c. 104 (Brownsey & Chinnock 1987). We therefore recognise it here as a distinct species, rather than as a subspecies of H. rugosula as proposed by Schwartsburd & Prado (2014).