Class
Subclass
Order
Family
Genus
Rhizomes long-creeping, 2–3 mm diameter, with stipes arising 20–35 mm apart; bearing multicellular, non-glandular, red-brown hairs up to 3 mm long. Fronds 240–1000 mm long. Stipes 70–450 mm long, 1–3.5 mm diameter, dark red-brown, bearing abundant colourless or pale brown glandular hairs up to 0.5 mm long and scattered red-brown non-glandular hairs up to 2 mm long. Rachises dark red-brown proximally, becoming paler and often green distally, densely covered in colourless or pale brown glandular hairs up to 1 mm long and scattered red-brown non-glandular hairs up to 2 mm long. Laminae 2-pinnate-pinnatifid to almost 3-pinnate, narrowly ovate to ovate, tapering to a pinnatifid apex, 130–550 mm long, 72–430 mm wide, dark green adaxially, lighter green abaxially, herbaceous; colourless and pale brown glandular and non-glandular hairs densely covering abaxial lamina surface and costae, 0.3-1.0 mm long on laminae, up to 1.5 mm long on costae; non-glandular hairs only on the lamina margins. Primary pinnae in 15–30 pairs below pinnatifid apex, widely spaced proximally, almost overlapping distally, the proximal pair arising at 30–70° to rachis; distal primary pinnae narrowly oblong or narrowly ovate; proximal primary pinnae narrowly ovate or ovate; the longest at or near the base, 37–240 mm long, 18–140 mm wide, apices acute or acuminate, bases short-stalked; costae winged distally. Secondary pinnae gradually decreasing in length along each primary pinna to the distal end; the longest ovate or oblong, 5–75 mm long, 4–26 mm wide, apices obtuse, bases short-stalked or sessile or adnate; costae winged throughout. Tertiary segments oblong, 3–14 mm long, 2–17 mm wide, apices obtuse, bases adnate, deeply divided on largest fronds. Veins ending in apices of ultimate segments. Sori ± round, protected by slightly reflexed green lamina flaps bearing a few short acicular hairs; paraphyses absent. Mean spore size 31–34 μm long, 20–22 μm wide; perispores pale, echinate and reticulate.
Hypolepis amaurorhachis is one of three similar species in New Zealand that have glandular hairs, red- or purple-brown stipes and rachises, and lack indusial flaps. It is distinguished from H. rufobarbata by the presence of colourless glandular and non-glandular hairs on the lamina margin in contrast to the abundant red-brown acicular hairs in H. rufobarbata. It also has colourless glandular hairs on the rachises and costae, whereas H. rufobarbata has red-brown glandular or acicular hairs. Hypolepis amaurorhachis is more similar to H. lactea but is distinguished by its paler red-brown stipe and rachis, becoming green in the distal half of the rachis, and by its longer glandular hairs on the laminae (0.3–0.7 cf. 0.1–0.3 mm long). Hypolepis amaurorhachis occurs only in the far south of the South Island and on the Chatham Islands and subantarctic islands, whereas H. lactea occurs in the North Island and South Island except for the far south. Hypolepis amaurorhachis can also be confused with H. ambigua, but is usually distinguished by the presence of glandular hairs and smaller spores (31–34 × 20–22 μm cf. 36–41 × 22–28 μm).
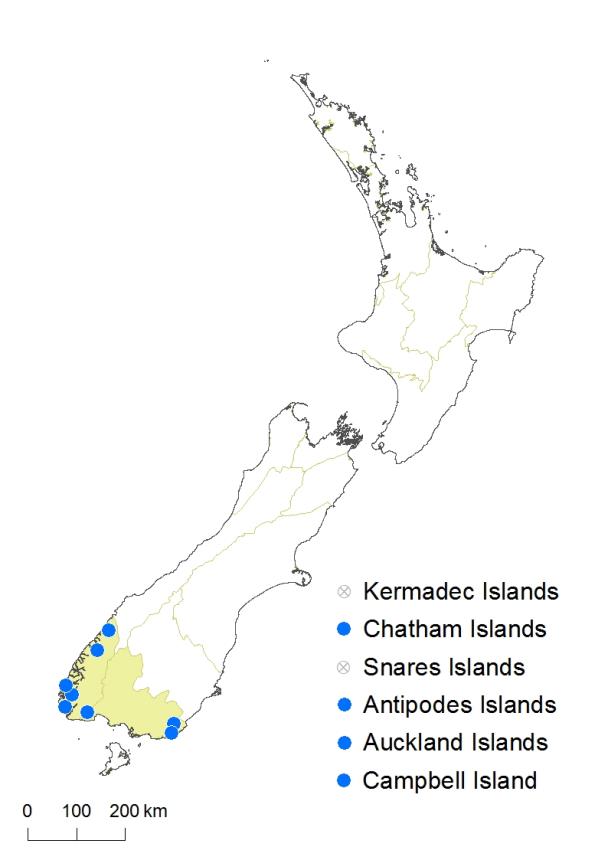
South Island: Southland, Fiordland.
Chatham Islands, Antipodes Island, Auckland Islands, Campbell Island.
Altitudinal range: 0–200 m.
Hypolepis amaurorhachis is known only from isolated collections in Fiordland and the Catlins region of the South Island, as well as from the Chatham Islands, Antipodes Island, Auckland Islands and Campbell Island. It ranges from near sea level in the South Island to 200 m on Campbell Island.
Also Australia (Victoria, Tasmania).
Grows on open rocky slopes and terraces near the coast, in open Dracophyllum scrub, in Bulbinella herbfield, or on tracksides and riverbanks, and in light gaps under podocarp or rātā forest.
Hypolepis amaurorhachis was given a conservation status of Naturally Uncommon by de Lange et al. (2018).
n = 52 (Brownsey & Chinnock 1984).
In his original description of this species in Cheilanthes, Kunze (1850) used the epithet "amaurorhachis". When Hooker (1851–1858) made the new combination in Hypolepis he spelled the epithet "amaurorachis", incorrectly citing the spelling used by Kunze. This error was followed by Brownsey & Chinnock (1987) when reinstating the name for the Australasian species, and perpetuated by subsequent authors (e.g. Brownsey & Smith-Dodsworth 2000). The original spelling used by Kunze is not an orthographic error and must therefore be retained as Hypolepis amaurorhachis.
In New Zealand, this species has been widely misidentified as Polypodium viscidum Roxb., Polypodium rugosulum Labill., Polypodium punctatum Thunb., or combinations based on them.
Hypolepis amaurorhachis occurs in both New Zealand and Australia, and, with n = 52, is cytologically distinct from the Australian H. rugosula, which has n = c. 104 (Brownsey & Chinnock 1987). We therefore recognise it here as a distinct species, rather than as a subspecies of H. rugosula as proposed by Schwartsburd & Prado (2014).