Class
Subclass
Family
Genus
Epiphytic, terrestrial or rupestral ferns. Rhizomes long-creeping, 0.5–0.75 mm diameter, bearing sparse to abundant pale brown hairs up to 1.2 mm long. Fronds 50–440 mm long. Stipes 12–90 mm long, dark brown or black, narrowly winged for most of their length, glabrous or sparsely covered in pale brown hairs proximally. Laminae usually 4-pinnatifid or rarely 3-pinnatifid, ovate or elliptic, 30–360 mm long, 20–130 mm wide, dark green, membranous, glabrous or with very scattered hairs on the costae. Rachises narrowly winged throughout, dark brown or black, often zig-zagged distally, glabrous or bearing a few scattered hairs proximally; rachis wings planate or slightly flexuose. Primary pinnae in 8–25 pairs, overlapping, winged throughout; distal portion of primary pinnae straight or slightly incurved acroscopically; distal primary pinnae ovate or elliptic or obovate, adnate; proximal primary pinnae narrowly ovate or ovate, adnate; the longest primary pinnae at or below the middle, 12–170 mm long, 10–20 mm wide. Secondary pinnae arising both acroscopically and basiscopically, scarcely overlapping, winged throughout, adnate; ovate or elliptic or obovate on distal primary pinnae, ovate or elliptic on proximal primary pinnae; the longest secondary pinnae 6–16 mm long, 6–10 mm wide. Ultimate lamina segments oblong, up to 5 mm long, 0.8–1.5 mm wide; apices obtuse or truncate; margins entire, lacking a distinct border; distal segments on primary pinnae divergent. Sori borne on short acroscopic and basiscopic segments throughout the pinnae, solitary, many on each primary pinna, adnate; indusia bivalvate; indusial flaps elliptic or orbicular or broader than long, 1–2 mm long, apices obtuse, margins entire, crested on outer surfaces; receptacles included within indusial flaps.
Hymenophyllum sanguinolentum is a small to medium-sized filmy fern distinguished by its entire lamina margins, black zig-zagged rachis, winged rachis and stipe, and crests on the outer surfaces of the indusial flaps. It is more or less glabrous throughout much of its range but generally somewhat hairier in the northern part of its distribution. When dry it has a strong smell and stains paper dark brown or yellow. It is a very drought resistant filmy fern, curling into a tight ball in dry weather. It is very similar to H. villosum, but is usually distinguished by its glabrous to sparsely hairy fronds, broader ultimate segments (0.8–1.5 mm wide cf. 0.4–1.2 mm wide), and indusial flaps which are often broader than long and have crests on their outer surfaces. The crests on the indusial flaps are not found in any other New Zealand species, but they are not always obvious, especially in dried specimens. Hymenophyllum sanguinolentum generally grows at lower altitudes than H. villosum (0–900 vs 370–1300 m in the North Island; 0–850 vs 30–1650 m in the South Island). Nevertheless there is a significant zone of overlap and plants of both species occur together in several places.
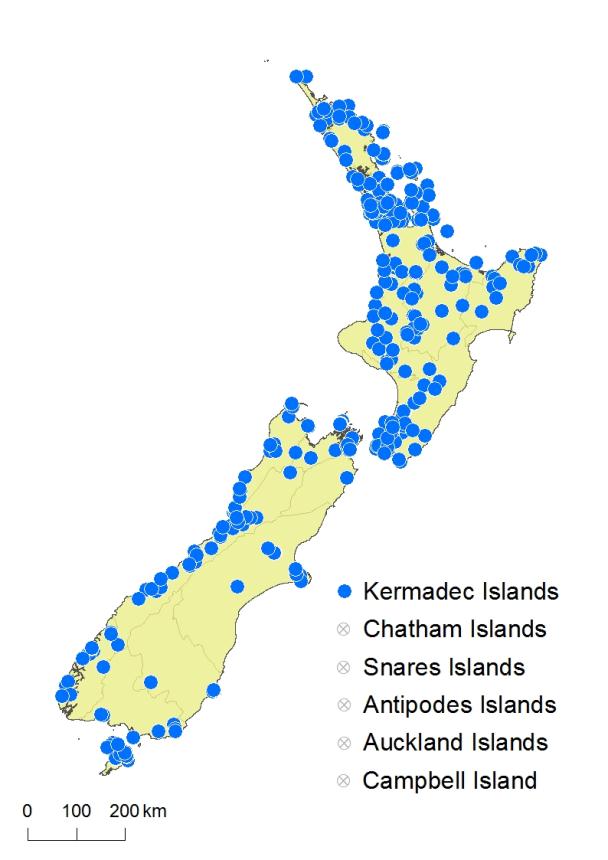
North Island: Northland, Auckland, Volcanic Plateau, Gisborne, Taranaki, Southern North Island.
South Island: Western Nelson, Sounds-Nelson, Marlborough, Westland, Canterbury, Otago, Southland, Fiordland.
Kermadec Islands, Stewart Island.
Altitudinal range: 0–900 m.
Hymenophyllum sanguinolentum occurs in lowland to montane forest throughout the North Island from Te Paki southwards, ranging from sea level to 900 m on Mt Honokawa, Raukūmara Range. In the South Island it mostly occupies lowland regions, principally west of the main divide but also in scattered localities in eastern areas, reaching 850 m on Banks Peninsula. It is apparently absent from much of central Otago and south Canterbury.
Also Cook Islands (AK 320150).
Occurs under kauri, podocarp, beech and broadleaved forest or in mānuka and kānuka scrub, growing on the ground, on rocks and lava, on banks, rock faces, rotten logs and stumps or as an epiphyte. It has been recorded on a wide variety of forest trees including Cyathea dealbata, C. medullaris, C. milnei, Beilschmiedia tawa, Carpodetus serratus, Dacrycarpus dacrydioides, Dacrydium cupressinum, Dysoxylum spectabile, Griselinia littoralis, Knightia excelsa, Kunzea spp., Leptospermum scoparium, Metrosideros excelsa, M. umbellata, Phyllocladus trichomanoides, Podocarpus totara, Prumnopitys ferruginea, P. taxifolia, Vitex lucens and Weinmannia racemosa as well as hanging from clumps of Astelia.
n = 34, 68 (Daellenbach 1982), 36, 72 (Brownlie 1954, 1961)
The name Hymenophyllum polyanthos (Sw.) Sw. has been widely misapplied to this species.
Hymenophyllum sanguinolentum was recorded from the Auckland Islands by Johnson & Campbell (1975) but the voucher specimen (OTA 33363) was misidentified and is actually H. villosum. It was also recorded from the Three Kings Islands, in error for the Kermadec Islands, by Brownsey & Smith-Dodsworth (2000).
Hymenophyllum lophocarpum was described by Colenso (1885) from collections he made in Seventy-mile Bush, County of Waipawa, from 1860–84. There are two specimens at K labelled "Hymenophyllum lophocarpum Col. sp. nov." in Colenso’s writing; one is annotated "New Zealand, W. Colenso, rec’d xii 1885" (photo WELT E468/32), and the other "70 m[ile] Bush, com. W. Colenso 5/1890" (photo WELT E468/31). There are another two specimens at WELT. One is labelled "Dannevirke, type of H. lophocarpum" in Cheeseman’s writing and "Hymenophyllum lopho/, sent Kew the best spns" in Colenso’s writing (WELT P002923). The other has a label copied from WELT P002923 in Zotov’s writing (WELT P003302). There are also two specimens at AK labelled "Dannevirke, type of H. lophocarpum" in Cheeseman’s writing (AK 83, 218893). Allan (1961, p. 26) cited a Colenso collection at WELT as the type but did not distinguish between the two sheets held there. However, Zotov’s label on WELT P003302 indicates that the material on that sheet is part of the collection labelled by both Colenso and Cheeseman (WELT P002923). It seems that Zotov segregated specimens from some of Colenso’s collections and re-labelled them (Brownsey 1979). In this case, Colenso’s material is now mounted on two sheets (with different numbers) but is part of the same specimen (Art. 8.3). Allan’s statement that the type is in WELT can therefore be taken as a lectotypification and relates to these two sheets (WELT P002923 and P003302).
Some plants of H. sanguinolentum and all of H. villosum that have been examined have n = 34 or 36. However, tetraploid plants of H. sanguinolentum with n = 68 or 72 have been recorded from the Port Hills, Banks Peninsula, Mt Grey and Peel Forest in Canterbury (see Dawson et al. 2000). Hybrid plants have also been detected at Mt Grey, Canterbury that show formation of trivalents, bivalents and univalents at meiosis (Lovis in Dawson et al. 2000). Analysis of the meitotic pairing indicated that these are likely to have been hybrids between tetraploid H. sanguinolentum and diploid H. villosum.
Some plants appear morphologically intermediate between the two presumed diploid species. Most frequently, such plants are recognised by having hairy fronds similar to H. villosum but indusia which bear crests on the outer surfaces similar to H. sanguinolentum. Plants of this description have been recorded from the Ruahine Forest Park (WELT P018742), Kaimanawa Forest Park (WELT P018741), Kaituna Valley, near Blenheim (CHR 46297), Kekerengu Range, Marlborough (CHR 387557), Blue Duck Reserve, Marlborough (CHR 387980), Port Hills, Christchurch (CHR 235775, 278448, 397939), Banks Peninsula (AK 278431, CHR 178338, 517421), Peel Forest (CHR 386696, 386702) and Tautuku Forest, Southland (CHR 403680). More rarely, plants lacking crests and having almost glabrous fronds have been recorded from Banks Peninsula (CHR 276234, WELT P000506, P005992), Alton Burn, Southland (CHR 403717) and near Dunedin (CHR 144311).
It is possible that the plants of intermediate morphology correspond to the tetraploid forms of H. sanguinolentum. The voucher specimens from the Port Hills for Brownlie’s report of n = 72 (CHR 397939), and from near Akaroa for Large’s unpublished count of n = 72 (AK 278431), are both listed above. However, further work is needed to determine whether all plants of intermediate morphology are tetraploids. If that could be demonstrated, it is likely that they represent an undescribed allopolyploid species derived from hybridisation between diploid plants of H. sanguinolentum and H. villosum. Apart from two records in the Ruahine and Kaimanawa Ranges, all plants of intermediate morphology have been collected from the eastern or southern South Island.