Class
Family
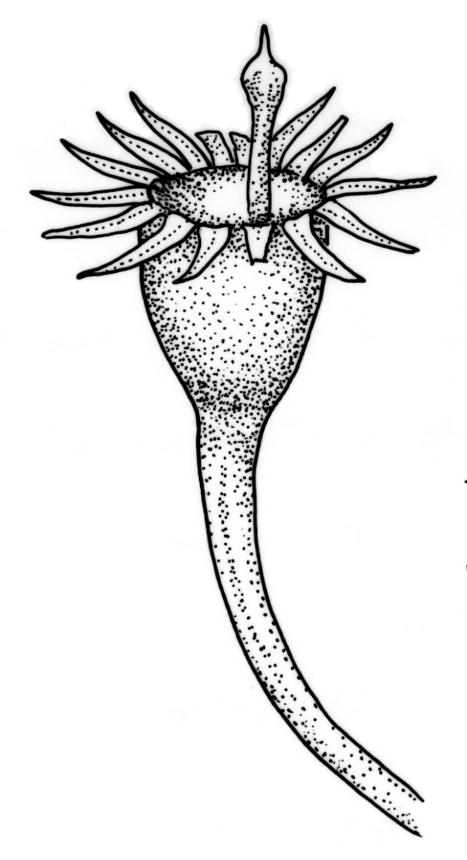
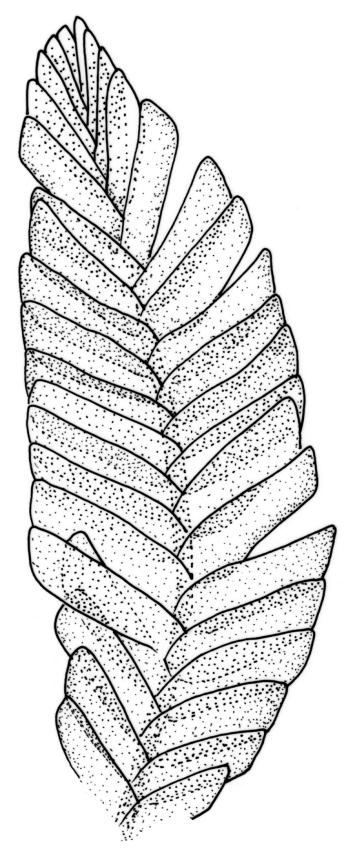
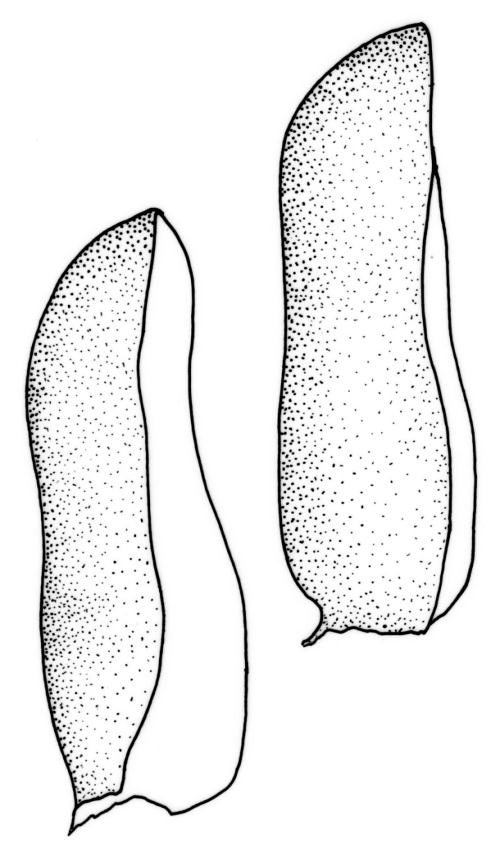
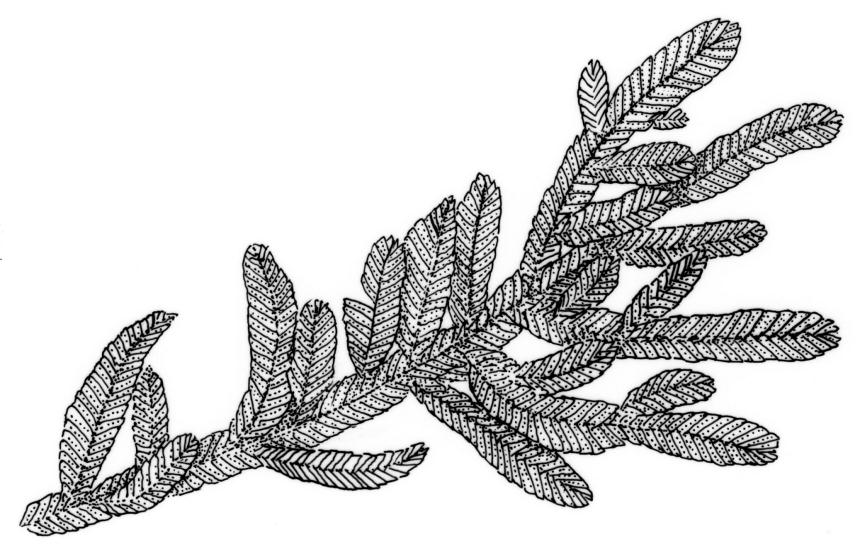
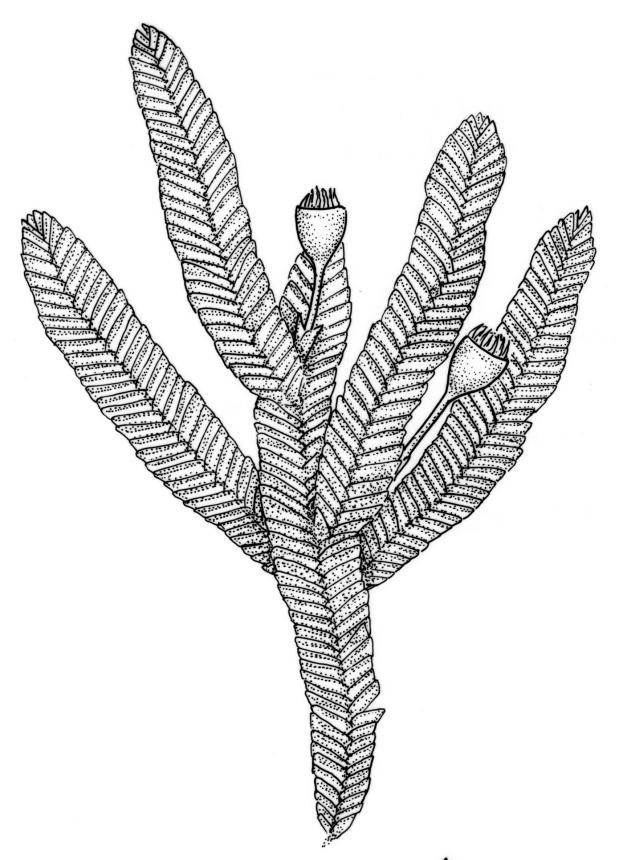
Plants small to medium-sized, strongly lustrous fresh or dry, yellow-green (rarely becoming brown or chestnut), strongly flattened, forming flat mats on bark or rock. Stems mostly to 25(–40) mm, irregularly to subpinnately branched, with scattered fascicles of pale brown, smooth rhizoids, in cross-section lacking a central strand and with the cells gradually becoming smaller at the periphery. Shoots c. 1.5–2.0 mm wide, broadly obtuse at apex. Branches mostly simple and c. 3–5 mm long, often denuded below due to deciduous leaves. Stem and branch leaves similar, distichous, closely imbricate, with an internal angle between the stem and the leaf of c. 60 degrees, becoming more nearly parallel to the stem near apex to form a broadly obtuse shoot apex, often deciduous below, symmetrically conduplicate, lingulate (under cover slip) with nearly parallel margins, rounded or truncate distally, strongly cucullate above, entire, mostly 0.85–1.1 × 0.3–0.35 mm (under cover slip); upper laminal cells linear, thin-walled, c. 60–72 × 4 μm, smooth; median cells of the cucullate leaf apex differentiated and forming a band c. 4–8 cells wide and extending to apex, quadrate or irregular, 9–15 μm in greater dimension (but appearing as a marginal band due to leaf folding); basal laminal cells shorter and more irregular; alar cells not differentiated. Costa absent. Axillary hairs concolourous with leaf, apparently one per leaf corner, composed of c. 5 uniformly oblong and smooth cells, c. 140 μm long. Pseudoparaphyllia absent.
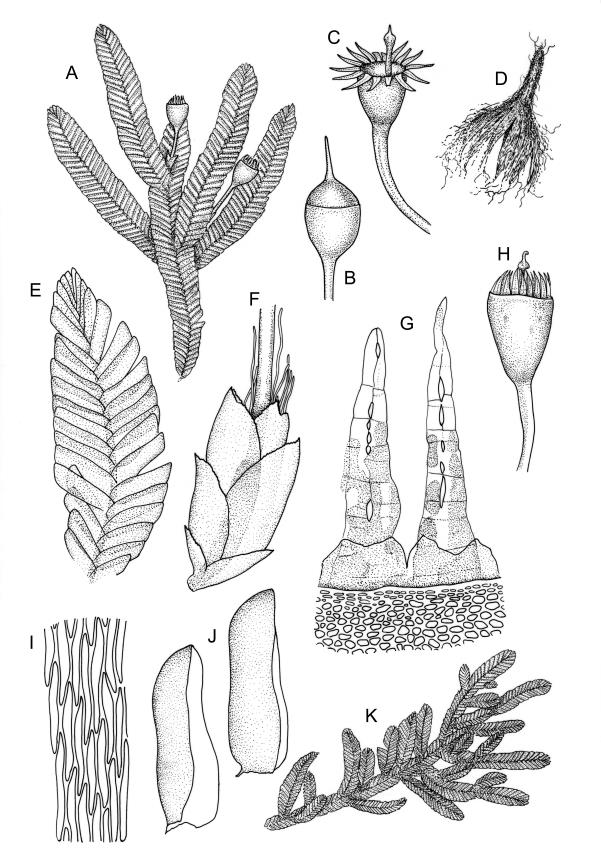
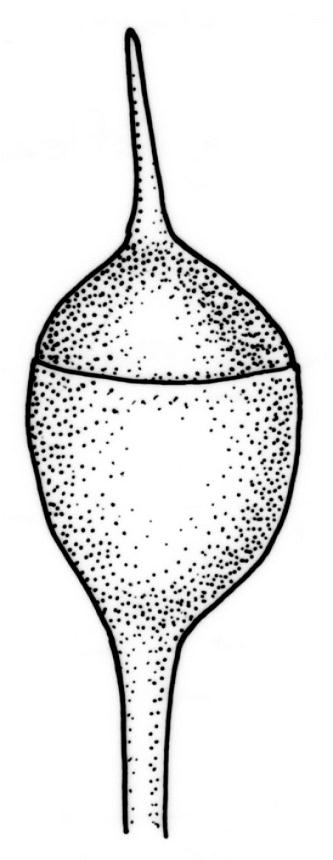
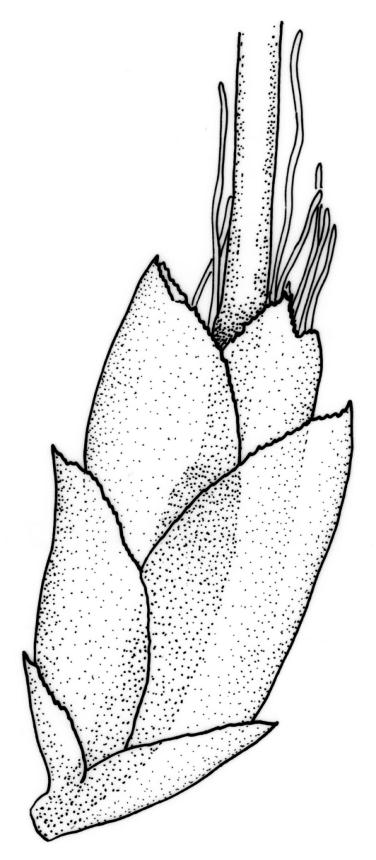
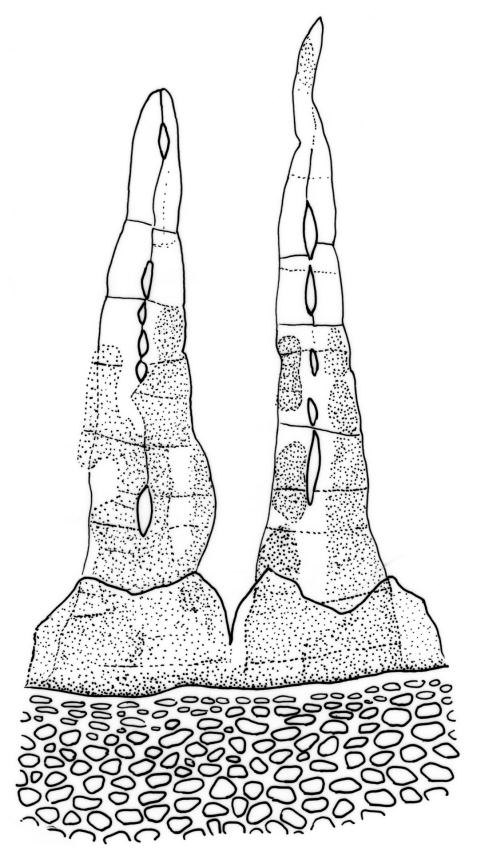
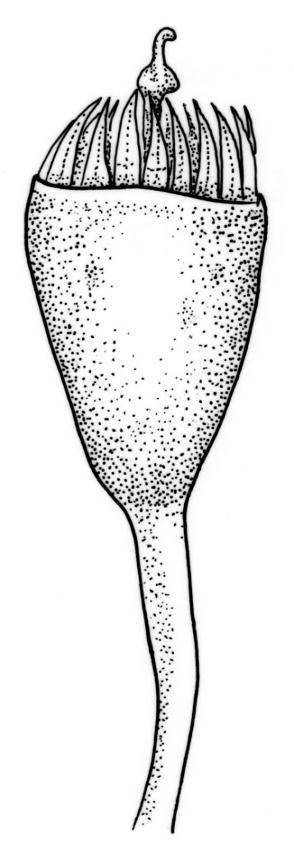
Reportedly dioicous. Perichaetia in axils of vegetative leaves; perichaetial leaves clasping, concave to ± conduplicate, irregularly laciniate at margins, apparently enlarging and sheathing the lower ¼ to ½ of the seta, enclosing c. 10 archegonia and several very long paraphyses which extend beyond the leaf apices. Perigonia not seen. Setae stout, red, straight, and smooth, 2–4 mm; capsules campanulate to cupulate, widest at mouth, with columella persistent and extending beyond the mouth, pale red-brown, 0.85–1.0 × c. 0.9 mm, lacking a differentiated neck; exothecial cells ± isodiametric, collenchymatous, mostly c. 20–25 μm diam.; stomata apparently absent; annulus not differentiated; operculum conic with an elongate and erect rostrum, ± equal the capsule in length. Peristome present, single in N.Z. material; exostome teeth widely spreading when dry, pale brown, smooth or nearly so, c. 325–375(–450) × 100–120 μm (but often shorter and irregular in outline), inserted c. 60–75 μm below rim, with a nearly straight divisural line, often irregularly perforate; endostome absent in N.Z. material; preperistome well-developed and obscuring the base of the teeth. Calyptra mitrate, deeply laciniate at base, hairy, enclosing the entire capsule and clasping at base. Spores 24–30 μm, smooth.
Lin 1983, pl. 5; Norris & Koponen 1987, figs. 7 e–i; Beever et al. 1992 fig. 60.
Orthorrhynchium elegans is most likely to be confused with Pseudotaxiphyllum falcifolium. By contrast P. falcifolium has leaves inserted in four rows and very strongly complanate. The individual leaves of P. falcifolium have markedly unequal conduplicate portions with the basal margin strongly inflexed (with the inflexed portion c. ⅓ the width of the remainder of the leaf and extending c. ⅔ the leaf length). In outline the leaves are oblong, but have a strongly asymmetric apex which is recurved and tapers rapidly to an obtuse apex; the leaf length is c. 1.7–1.8 mm (compared to c. 1.0 mm in O. elegans); the shoots are typically c. 2.8–3.0 mm wide (compared to c. 1.5–2.0 mm in O. elegans). The elongate setae (c. 14–20 mm) and the nodding, ovoid to oblong-ovoid capsules with a double peristome of P. falcifolium further distinguish it from O. elegans. Pseudotaxiphyllum falcifolium is predominantly a terrestrial species whereas O. elegans is predominantly epiphytic, although both species can occur on limestone.
K; NI: N Auckland, including offshore islands (HC, LB, GB), S Auckland, Gisborne (Raukokore River, Whinray Scenic Reserve, Rākauroa Scenic Reserve, Mangapōike Falls), Hawke’s Bay (Kapokaponui, Sentry Box Scenic Reserve, Mohi Bush Scenic Reserve), Wellington; SI: Nelson, Marlborough, Canterbury (Wainui), Westland (Greymouth), Otago (eastern regions, including near Dunedin), Southland; St; Ch. Orthorrhynchium elegans is a frequent species in much of the northern NI and it is often collected through most of Gisborne and Wellington L.D. Herbarium holdings of this species are inexplicably uneven in distribution and it seems genuinely uncommon in many regions including Hawke’s Bay, Taranaki (no confirmed localities), Canterbury, and Westland L.D. and all but the eastern portions of Otago L.D. In her checklist of Banks Peninsula mosses, Macmillan (1996) recorded one early R. Brown collection from Wainui, and this remains the sole confirmed Canterbury L.D. record.
Anomalous. Mainland Australia* (N.S.W., Qld), Norfolk I.*. Sri Lanka*, and New Guinea* (confirmed from P.N.G.; reported from West Irian). Lin (1983. p. 78) recorded O. elegans s.l. also from India and Lord Howe I.
Epiphytic on a very wide range of dicotyledonous trees including Ascarina lucida, Beilschmiedia tawa, Carpodetus serratus, Coprosma spp., Corynocarpus laevigatus, Dysoxylum spectabile, Elaeocarpus hookerianus, Fuscospora solandri, Geniostoma sp., Knightia excelsa, Laurelia novae-zelandiae, Lophomyrtus bullata, Melicytus ramiflorus, Myrsine spp., Pennantia corymbosa, Pseudopanax spp., Syzygium maire, Vitex lucens, Weinmannia racemosa, and W. silvicola; the native conifers Agathis australis, Dacrycarpus dacrydioides, Dacrydium cupressinum, and Prumnopitys ferruginea; as well as the introduced Cupressus macrocarpa. Vitex lucens is the most frequent host tree species in N Auckland L.D. (Jessica Beever, pers. comm.). Also occurring on dry, shaded limestone. One collection from D’Urville I. (Marlborough L.D.; P.J. Brownsey s.n., WELT M010275) was made from ultramafic rock. This species most often forms pure or near pure mats, although Cyrtopus setosus is an occasional associate. On North I. occurring from near sea level to at least 750 m (Erua, Wellington L.D.) and on South I. to at least 620 m (Mt Burnett, Nelson L.D.).
In a N.Z. context O. elegans is recognisable at a glance by its epiphytic habit and very lustrous, strongly distichous, and conduplicate leaves. The leaves diverge from both the stem and branches with an internal angle of c. 60 degrees. The tightly imbricate and uniformly-sized leaves form what Streimann (2002, p. 121) aptly described as "a continuous neat straight edge." At the shoot apices the internal angle becomes smaller (the leaves becoming more nearly parallel to the axis of the branch), which produces a characteristic broadly obtuse shoot apex. In some populations, the leaves of the lower portion of the shoots are deciduous and only leaves at the shoot apex persist. The leaves are so strongly imbricate that the leaves cohere even after they have been stripped from their stem or branch and the conduplicate leaves cannot be flattened without tearing. The conduplicate portions of the leaf are equal in width, which helps distinguish O. elegans from some superficially similar species (e.g., Pseudotaxiphyllum falcifolium and the tropical Pacific Cryptogonium phyllogonioides, discussed above). The differentiated band of median cells in the leaf apex (the "cucullus" sensu Lin 1983, p. 80) is visible under the compound microscope.
Despite repeated efforts, I have been unable to locate perigonia in this species and have relied on Lin’s (1983) statement that the species is dioicous. The rarely-seen campanulate or obovoid capsules are on short setae and the lacerate and clasping calyptrae give this species further distinction. I have never seen an endostome here, although Norris & Koponen (1987) suggested that it can be present and "reduced to few remnants of cilia."